Halle & Olderman in the 1970s created a model of 23 types of architectural models to which all tree species are believed to conform. They started with the idea that the shoot apical meristem/s (“SAM” – the primary growing tip) is/are the ‘treemakers’.ref The behaviour of the SAM over time determines the form of the tree. They identified four types of meristems (active growing shoots) which led to different forms – those with a single meristem (like a palm), those with modular construction which follow a precisely repeating pattern, trees with trunk/branch differentiation and those in which the meristem changes direction to produce both trunk and branch.ref These depended on factors like whether the SAM is reproductive or vegetative, whether it grows vertically or horizontally, whether growth of the SAM is continuous or rhythmic (ie. has a period of dormancy or a growth pause) and the chronology of growth of each meristem.ref
Below is an updated version which includes some models added later (and one which was removed as nobody found a real tree which conformed to the theoretical model).
https://gali-izard.arch.ethz.ch/francis-halle
Each architectural model has a unique combination of growth, branching, axis differentiation and position of sexual structures.ref Not all of these are relevant to bonsai – as you can see Holttum & Corner don’t branch and Tomlinson & Bell branch underground. Many of these models are relevant for palms, cycads and tropical trees which aren’t commonly used for bonsai.
Two of the architectural models represent many of the trees used in bonsai at least in the Northern Hemisphereref – Massart’s and Rauh’s model – their main difference is in the branching angle with Massart’s at an angle to the stem and Rauh’s growing upwards. In both models the trunk is monopodial – it keeps extending upwards and is dominant.
Massart’s ModelRauh’s Model
Massart’s model represents Abies, Picea, Sequoia, Metasequoia, Cedrus, Taxodium, Taxus, Cephalotaxus, Ginkgo & Ilex aquifolium. More forms and discussion of Massart’s model are represented in this post.
Rauh’s model covers the Cupressaceae family (cypress, juniper & redwood), some Araucariaceae, the Pinaceae family including most Pinus species, the Podocarpaceae family, as well as angiosperms such as oak, maple and ash. This is shown in more detail in this post.
If you’re looking to understand the architectural model for your particular tree, you might consult this book – it mainly focuses on tropical trees but gives some pointers on working it out.
Some other models include Attim’s model for Eucalyptus, this is similar to Rauh’s model but follows a continuous growth pattern – in these trees as one leaf expands outside the bud it is replaced by a new bud initiated at the shoot apex.
Troll’s model is applicable for hemlock, acacia, beech, where “axes are all plagiotropic (ie. horizontal), the architecture being build by their continual superposition; main-line axes contribute part trunk, part branch, the proximal part becoming erected, most often secondarily after leaf fall” – it is believed that reaction wood is involved in determining this architecture (the type of wood created to stabilise a branch against gravity – compression wood developed under the branch in the case of gymnosperms, and tension wood developed above the branch in the case of angiosperms). Hemlock is a gymnosperm with this model.
Troll’s Model
Finally, trees can move from one model to another when they move from their juvenile vegetative phase to their reproductive phase. For example Apple has been found to conform to Rauh’s model when juvenile but Scarrone’s when reproductive:ref
The reproductive system and organs of plants are extremely varied and complex, and worthy of an entire website to themselves – a comprehensive view is beyond the scope of this website. But what I want to do is provide a bit of information to help you identify which buds might be reproductive vs vegetative.
There are different buds for vegetative (shoots & leaves) and reproductive (flowers, strobili) organs – within each bud a different set of components develop depending on what kind of bud it is. For angiosperms, it’s hypothesised that flower buds are based on the same structures as vegetative buds – that is, a bud starts as vegetative and then differentiates into a flower bud.ref For gymnosperms, reproductive buds contain male or female strobili.ref These are the male pollen cones or the female seed cones.
There are two ways to work out which bud is which – by their appearance or by their position on the tree. The appearance route is best aided by dissecting some actual buds from the tree you are interested in, so you have real data from the real tree. Otherwise, read on for more information about how vegetative and flower buds differ in appearance.
Vegetative buds are “encased by strong, coarse, mature scale leaves. Thinner, more membranous scale leaves make up the next layer. The scale leaves form a protective enclosure surrounding the developing foliage leaves.”ref In many of the articles online, leaf buds are said to be thinner than flower buds (at least for angiosperms). Below are the vegetative buds of Acer pseudoplatanus and Fraxinus excelsiorref:
A flower bud develops sepals, petals, stamen, pistil, ovaries & anthers, instead of leaves and more buds. Below is a scanning electron micrograph of a Bing cherry flower bud forming where M is the meristem, B is the bract and F is the very start of the flower forming – the progression over time is from left to right. On the right is the pistil with ovary (O) style (SY) and stigma (SM). The scale of the image is provided by the white bar on the bottom right hand side, which is 100 micrometres (or microns) – about 1/100th of a centimetre. Obviously it’s impossible to detect a flower bud at this stage by eye, it’s way too small!
The buds of these Bing cherries started forming and were detectable under a microscope from mid-May (the location was Washington state USA) – the year before they would flower and fruit. At a lower magnification below is a progression in flower bud development of a Camellia – the key difference is that at A2 when the flower starts to differentiate, the tip of the bud becomes more rounded and flattens.
Some of the trees we use in bonsai are dioecious which means they are only male or female and not bothref. This means they will produce only one type of flower or strobili bud. To check whether your gymnosperm is dioecious or monoecious you can check the gymnosperm database. Unfortunately I don’t have a reference link for angiosperms.
In gymnosperms there are no flowers, instead the reproductive organs are male or female strobili (pollen or seed cones respectively). A brilliant reference for some of these is available online here. An extract from the publication is provided below with some examples of vegetative and male & female strobili buds.
As noted another way to determine the type of bud is by its position. To do this you need to work out what the architecture of your tree is and where buds of different types typically form. Trees conform to one of 24 architectures as described in Tree Architectural Models and these give an indication to the location of reproductive buds. Trees can have one model for their juvenile phase (before they flower or adopt mature foliage) and then one for their reproductive phase.
In the case of apple, it has been found to conform to Rauh’s model when juvenile and Scarrone’s when reproductive.ref Even with this, “great differences exist between cultivars whether they belong to “spur” vs. “spreading” or “terminal bearing” growth habit”ref
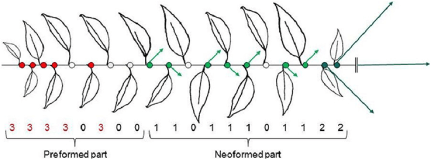
In cherry, “The long branches bear short shoots, also called spurs, in lateral positions on the distal half or two thirds of the branch with more vigorous spurs toward the distal part [the most distant part]. Flowering occurs in axillary positions on the five to six basal-most nodes of all shoots whether long or short. Floral buds are thus located exclusively on the preformed nodes of the previous year shoots.”ref
Red dots are flower buds, formed the previous year, and green dots are vegetative buds, formed during the growing season https://www.frontiersin.org/files/Articles/105157/fpls-05-00666-r2/image_m/fpls-05-00666-g001.jpg
In some species, particularly pines, the long shoot buds have many different components on them in a specific order. Here is a Pinus contorta bud – you can see the male (pollen) cones appear first, and are positioned at the bottom of the final shoot, and the female (seed) cones appear last just below the top of the shoot.
As you can tell from the above, there is quite a bit of complexity in understanding the architectural model of a tree, and where different buds form, as there are many variables involved. Observation is probably going to be the better method.
Most bonsai trees progress through stages of development, each with a different objective. In general the progression is thicken trunk -> achieve branch & root structure -> achieve branch, foliage & root ramification -> reduce leaf size -> evolve as branches grow/fall off. The faster we can move through the first few development stages, the faster we will have beautiful, well-proportioned bonsai – harnessing the tree’s natural growth is a way to speed this up. We also want to avoid doing things which slow down a tree’s growth during these phases, as this will mean it takes longer to get the tree we want. Read about how trees grow before starting at #1 below. Also consider what do old trees look like?
1. Trunk
Some bonsai enthusiasts collect mature trees for bonsai specifically so they can start with a thick trunk, following a collection process which minimises damage to the tree. The alternative is growing your tree’s trunk. Once a tree has its roots and foliage reduced in size in a bonsai pot, it won’t generate the energy needed to make significant sapwood additions and its girth will only increase by small increments every year. So you really need to be happy with the trunk size first before you stick it in a tiny pot. But – how big should a bonsai tree’s trunk be?
2A. Branch Structure & Overall Shape
Arranging the branches is what gives you the canopy and overall foliage shape that you’re after and the first step in this process is growing (or developing) the branches you want in the positions they are needed. Growing a branch starts with a new bud, which, unless it’s a flower bud, becomes an extending shoot and eventually a new branch. So firstly you need to work out where new buds will grow on your tree and then deal with the extending shoots as needed to get the required internode length.
You may need to remove some buds and shoots if they don’t help achieve the shape you are looking for – this should be done as soon as possible to avoid wasting the tree’s finite energy reserves. You have a trade-off to make here because leaving more foliage on the tree will provide more energy overall which contributes to its health and ability to recover from interference. However, growing areas of the tree which won’t be part of the future design is a waste of energy. You don’t want to remove so much of the tree’s foliage that it struggles to stay alive or develop the areas that you do want to grow out.
When you are creating your branch structure, often you will need to reposition branches – this is done with a wide range of different tools and techniques. A more advanced technique for adding new branch structure is grafting.
Sometimes the trunk itself or larger branches need a rework, to make them more interesting or to make them look more like old trees – for example adding deadwood or hollowing out the trunk. Usually this is achieved through carving.
2B. Creating a Strong Root System
The trunk thickening and branch structure phases both work best when the tree has lots of energy and so letting it grow in the ground or in a decent sized pot during these phases will get you there quickest. This also allows the roots to keep growing, but you want to understand about the role of roots, and root structure & architecture even if you still have your bonsai in a training pot. Particularly in this case, knowing about how to foster the the rhizosphere will help your tree stay vigorous. To maximise the roots’ exposure to nutrients and water you want to encourage Ramification of Roots (lateral root development).
Eventually it’s time to move the tree into a bonsai pot. This requires cutting back the roots, but as long as the roots are balanced with the foliage in terms of biomass, the tree should be OK. Root growth is usually prioritised outside of times of stem/foliage growth, and above 6-9 degrees C. So repotting might be best conducted at times that meet this criteria. Your growing substrate/medium is an important consideration.
3. Ramifying Branches & Foliage
Ramification is when branches subdivide and branch, giving the impression of age and a full canopy – and a well-ramified tree is a bonsai enthusiast’s goal. There are some techniques for increasing the ramification of branches and foliage. But not as many as there are for root ramification.
This stage also involves ongoing branch selection and reshaping (see 2A above). Another consideration is whether to keep or remove flower buds.
4. Reducing Leaf Size
An end stage in the journey to bonsai perfection is leaf size reduction. In nature, leaf sizes reduce relative to the biomass of the tree as it ages but since bonsai are small this effect doesn’t translate since the biomass never gets large enough. The tried and tested method for reducing deciduous tree leaf size is actually to practice one of the various methods of defoliation. A couple of others are covered here in reducing leaf size.
When to conduct these various activities depends on when the tree can best recover from them – which is a function of the Tree Phenology (or Seasonal Cycles).
5. Evolving Branches
Trees are not static organisms – they obviously continue to grow which is what we harness in the above steps. Part of this is that eventually branches may become too large for the design, or they may fall off (Peter Warren notes that Mulberry are known for this). As bonsai artists we want to have this in mind so that branches are being developed which can take their place in the future. This is an ongoing version of step 2A.
Once your tree has grown in the general direction and shape you want, you can refine it through pruning. Cutting into a tree can affect its health & vigour, so it’s helpful to understand exactly what happens to a tree when you do this. A really excellent paper explaining the effect of pruning is available from Purdue Universityref but to summarise, pruning has the following effects on a tree:
it removes photosynthetic material (leaves) thereby reducing the tree’s ability to generate energy
it reduces transpiration (the evaporation of water from the canopy) and the rate of water transport up the tree
it disrupts the pathways of plant growth regulators, causing regrowth but also consuming stored energy
if the main xylem vessels in the trunk are cut, it causes embolisms which reduce the water carrying capacity of the tree
it exposes the internal vascular system to the environment where bacteria and fungi can enter (by causing a wound)
on some conifers, pruning the shoot or branch removes options for future bud growth because dormant buds and meristem tissue is often concentrated in the more recent growth
Minor Pruning
Minor or leaf pruning is used in bonsai to keep the shape of a tree according to a design, but also to create ramification and reduce leaf size (or, keep leaves small). As per point 3 above, pruning leaves drives the tree to refoliate and it does this by activating dormant or suppressed buds. In deciduous trees there is usually a bud in every leaf axil and this will go on to produce at least 2 shoots, so you also get increased ramification. With only stored reserves to use for refoliation, shared across twice as many buds, leaf size will be reduced. Read more in: ramification of branches and foliage.
Major Pruning
Major pruning which involves cutting off branches or significant parts of the foliage may have more impact on the tree. The first thing is that removing large amounts of foliage will reduce the tree’s ability to generate energy. It will also reduce the tree’s energy requirements but not by as much as is lost (since leaves are working for the whole tree and not just to sustain themselves). See this article: Defoliation.
Major pruning is often required to get the design you want for a bonsai. So is it better to grow out then cut back, or cut back then grow? Growing first generates lots of energy but also lots of wasted growth, which is eventually removed. Cutting first saves energy by directing it all to the places you want to develop on the tree, but it reduces the total amount of energy available for growth.
To test this look at the following calculation. If you start with two identical 50-leaved plants, and the goal of reaching a particular level of refined foliage in 5 years time, you have two options. Scenario 1 lets the plant grow unpruned all the way to the end of the period then has a major prune back down to the target level of foliage. Scenario 2 prunes every year, gradually building up to the target level. Although they start and end in the same place, the first plant has generated a whopping 195,250 ‘leaf units’ of energy for growth – 12x what the second plant has generated.
cut and grow model
As much as 80% of the energy created by leaves is exported to the other organs of the plantref. These energy units could have been used in places that don’t eventually get removed in the ‘Cut’ scenario, such as thickening the trunk, storing reserves for stronger budding or refoliation.
The most obvious risk with major pruning is the fact that you are effectively wounding your tree. Read more about how it responds in repairing (?) damage.
You’ve probably heard of rooting hormone powder, or auxin, or gibberellins – these are all ‘Plant Growth Regulators’. Plant Growth Regulators used to be known as ‘phytohormones’ which means plant hormones. This has been quite a contentious topic among plant biologists.
A hormone in an animal is a chemical messenger, a substance which acts as a signalling or control molecule to cause an action to take place. “Hormones carry out their functions by evoking responses from specific organs or tissues that are adapted to react to minute quantities of them”ref. In animals, hormones are produced at a specific site (a gland, like the pancreas), work at low concentrations, and have a predictable dose-response. That is, an increase in hormone will result in the more of the related action (eg. more insulin leads to more sugar being taken up by the liver).
There *are* substances synthesised by plants which are involved in regulating growth – plant growth regulators – but they don’t work in the same way as animal hormones. It’s said that the assumption that they did sidetracked plant researchers for decades.ref Plant ‘hormones’ are synthesised in multiple sites in a plant (and potentially in every cellref), have multiple actions on different cells (they don’t act on just one organ or tissue), don’t exhibit predictable dose-response behaviour like animal hormones and are involved in significant interactions or feedback loops with each other.ref
What this means is that it’s quite hard to unpick what they do and how they work. The roles and mechanisms of plant growth regulators are still very much current research topics, as can be seen at two of the research groups at Cambridge University’s Sainsbury labref1,ref2 Some of the early theories about them were comprehensively demolished in a seminal article by Anthony Trewavasref (in particular the theory of auxin-derived apical dominance which was later proven wrong as explained below).
We now know that plant growth regulators act in concert with genes and the proteins they express, and not as an independently acting substance (one of the genes involved in cytokinin synthesis is known as LONELY GUY…).
So what are the plant growth regulators, where and how are they made? There are nine main plant growth regulators you may come across in your reading:
Auxinref – classically called ‘the growth hormone’ and a signal for division, expansion, and differentiation throughout the plant life cycle – involved in root formation, branching, the tropic responses, fruit development, shoot meristem function, the development of cotyledons and senescence. The most common form is Indole-3-acetic acid (IAA). Auxin acts in a ‘ying-yang’ relationship with cytokinin (see below)ref as well as with gibberellins. More about auxin below.
Cytokinins (CK)ref1ref2 – involved in cell division, shoot initiation and growth (including maintaining the stem cell niche), nutritional signaling, root proliferation, phyllotaxis, vascular bundles, leaf senescence, branching and nodulation, seed germination, nutrient uptake, and biotic and abiotic stress responses. 6-BAP or 6-Benzylaminopurine is a synthetic cytokinin which is used in micropropagation and agriculture. Coconut water (not milk) has been found to be a natural source of cytokinins.ref
Brassinosteroidsref – involved in a wide spectrum of physiological effects, including promotion of cell elongation and division, enhancement of tracheary element differentiation, retardation of abscission, enhancement of gravitropic-induced bending, promotion of ethylene biosynthesis, and enhancement of stress resistance.
Gibberellins (GA)ref – involved in multiple processes including seed germination, stem elongation, leaf expansion and flower and fruit development.
Strigolactones (SL)ref – induce hyphal branching of arbuscular mycorrhizal fungi and are shoot branching inhibitors.
Abscisic Acid (ABA)ref – involved in the induction and maintenance of seed dormancy, stomatal closure, and response to biotic and abiotic stresses.
Jasmonates (JA)ref – shown to be inhibitors of growth but also involved in development of flowers and defense responses against herbivores and fungal pathogens
Salicylic Acid (SA)ref – associated with disease resistance
Ethyleneref – multiple effects on plant development including leaf and flower senescence, fruit ripening, leaf abscission, and root hair growth.
Slightly maddeningly none of these substances do just one thing – they’re all involved throughout the plant!
So how and where they are made in a plant? This isn’t simple either. In fact local biosynthesis is thought to be critical for plants, whereby plant growth regulators are made at the site where they are needed. For example both auxin and cytokinin are synthesised in leaves *and* in rootsref, and can be made by chloroplasts and mitochondria, organelles which occur throughout the plant.ref Chloroplasts can make precursors to auxin, abscisic acid, jasmonates and salicylic acid.ref
Even though plant growth regulators don’t act in a predictable dose-response way like animal hormones, they still have a role in shaping plant growth in tissues which are sensitised to respond to them. Theoretically by understanding these responses we can manipulate a tree’s growth. And this is what they do in plant tissue culture (more on that below).
You may have heard the theory of auxin-controlled ‘apical dominance’, which holds that auxin produced by leaves at the apex inhibits lateral buds. This theory was strongly criticised by Trewavas in 1981: “The only hypothesis of apical dominance which has retained some measure of support is the nutritional one. A number of plants placed under conditions of reduced nutritional status adopt a growth pattern of strict apical dominance.” His point of view was further supported by a 2014 study which found that “sugar demand, not auxin, is the initial regulator of apical dominance”.ref The researchers found that after removal of the shoot tip, sugars were rapidly redistributed over large distances and accumulated in axillary buds within a timeframe that correlated with those buds releasing. But auxin didn’t travel fast enough to be responsible for bud release. So basically they found that apical dominance arises because the main shoot is greedy for sugars, and due to its position at the end of the vascular system it can prevent lateral buds from taking the sugars needed to release and grow.
Auxin does play some role though, and the theory is that its role related to the fact that it’s the only plant growth regulator which displays polar transport. That is – it moves from the apex to the base of the plant, via the phloem, and can travel the entire length of the plant, ending up in the roots. This gives auxin a special role related to the spatial aspects of growth, and auxin ‘maxima’ (locations where auxin accumulates) are sites where new buds, flowers or lateral roots emerge. In fact, auxin and cytokinin work in concert throughout the plant, from shoots to roots, with apparently opposite effects in each location “like yin and yang”.ref
An excellent reference point for this subject is the world of plant tissue culturing. This is where small pieces of plant tissue are sterilised and cultured in a medium containing plant growth regulators, which cause the tissue to grow into a ‘plantlet’ (sometimes in a test tube, if the source material is small). Further steps multiply the plantlet into several plantlets, which are then encouraged to create roots, multiplied again and/or planted out as seedlings to harden off. This process is used for industrial plant cloning where large numbers are required, and in the aquarium trade to avoid contamination with snails and other microbes (see Tropica’s website).
In plant tissue culturing, plant growth regulators are used to induce the relevant growth stage, which ones work for each species in which stage is documented in the ‘protocol’ for that species. In all cases specific ratios of cytokinin:auxin (and sometimes gibberellins) lead to different developmental stages – shoot growth, lateral shoot growth and root growth.ref1, ref2 To give you a bonsai-oriented example, one study determined a protocol for the micropropagation of Prunus Mumeref. They were able to multiply fresh prunus mume shoots in a petri dish using a 4:1 ratio of cytokinins to auxins, and then root them using auxin.
So – apologies for the rather long read, it is quite a complicated subject! What can we take from all this for our bonsai practice? Firstly we can stop the brain-bending trying to understand how auxin controls apical dominance because it doesn’t – access to sugars does this instead.
Also we can use the yin-yang rule – high cytokinin:auxin encourages buds & shoots, and high auxin:cytokinin encourages roots. So I’m going to start adding some auxin rooting gel into my air layers and cuttings. Cuttings have never worked for me in the past so maybe this will be the secret sauce I need. I’m also going to try some cytokinin gel to encourage lateral budding on my trees.
If you are looking for products to give this a try, make sure the product actually contains the plant growth regulator you want. For example, Clonex contain auxin, and some of the orchid budding pastes such as Keiki paste contain Kinetin (a cytokinin). Many other ‘rooting hormones’ or plant hormones products on the market have no ingredient list at all, so avoid those. You can also find these products online in shops dedicated to hydroponics, where cloning and plant tissue culturing is a technique used by practitioners, or in lab supply shops such as microscience or Phillip Harris in the UK. You can even make your own hormone gels following these instructions.
Another trick you can use is that gibberellic acid can be used to break dormancy in seeds, if you really don’t have the patience to wait for natural dormancy to break. Or give coconut water a try, this has been found to have a similar effect in a range of species. For hard coated seeds in particular, usually it’s best to search Google Scholar for a researcher who has experimented with different approaches, since what works is very species-dependent.
Gymnosperms relevant for bonsai include ginkgo and the Pinales order (Araucariaceae, Cephalotaxaceae, Cupressaceae, Pinaceae, Phyllocladaceae, Podocarpaceae, Sciadopityaceae & Taxaceae – this is explained in The kingdom Plantae and where trees fit in). Ginkgo is a special case described separately at the end of this post.
So what we’re interested in in bonsai is where lateral buds appear, and in particular whether they can develop adventitiously (or backbud). Angiosperms (flowering plants) are relatively easy to understand in terms of their lateral budding, as many species reliably produce a bud in each leaf axil (the axil is the place on the stem where the leaf is/was connected). In gymnosperms though, this is not as predictable and it’s not the case that each needle contains a bud – at least not in every species and not detectably. And looking at the different foliage forms below, you can see that different bud types must be involved to generate all these different leaf models.
Many conifers have a terminal bud at the end of each long shoot/branch surrounded by a number of close lateral buds in what’s called a ‘whorl’. These include pines, spruce, fir, and the Auracaria family. The whorl in the picture is a Scot’s Pine, with a vegetative bud in the middle and reproductive buds around it. This will usually be the apical or strongest bud, receiving the majority of the sugars from photosynthesis.
When the vegetative bud extends, it is called a ‘candle’ because it is a long thin structure – which looks like a candle. Below you can see a Pinus Thunbergii (Japanese Black Pine). Some candles are extending and some have extended and formed cones from the lateral buds around the main bud. No branching will occur from reproductive buds as they terminate the shoot.
https://www.conifers.org/pi/pi/t/thunbergii02.jpg
Bonsai enthusiasts commonly prune the candles to maintain a short needle length, this has the effect of arresting the needle growth; it is also possible to completely remove the candle, to force bud break at the base of the candle which results in smaller and more buds. In pines there are usually short shoot buds at the base of the candle – these will produce needle clusters in the future but no stem elongation. Breaking or pruning the top of the candle will activate these buds, which is good for ramification. If you want to continue developing the structure of the tree, you need a long shoot with a terminal vegetative bud as this won’t fall off.
As well as the terminal buds, pines sometimes have buds on their lateral shoots, between the needles, as well as internodal buds, which appear along the stem and not just at the end. These usually appear at the axil of the individual leaves on a long shoot/stem (Dörken, 2012).
Other conifers such as those in the Cupressaceae family (Thuja, Juniperus, Cypress) do not have whorls or needles, they have scale-type leaves in ‘branchlets’ (and needle-like leaves when juvenile). You can see below some examples of these which show the lateral buds forming from inside the lateral leaves (the leaves on the sides of the shoots). Since these branchlets squeeze a lot more leaves in, they have more potential for budding than do individually-leaved species such as Abies (fir) and Picea (spruce).
However one key attribute of species in Cupressaceae like these scale-leaved ones above is that just like pines they do still have differentiated short shoots and long shoots (Dörken, 2012). The short shoots are the individual branchlets, which abscise as a unit after a few years (detach from the long shoot and fall off). At the base of this short shoot is another bud waiting to generate a new shoot once the branchlet falls off. So new foliage will come from the leaves on the branchlet while it is active, and then from where the branchlet was connected to the stem when the whole branchlet falls off.
Conifers with individual needles such as firs and spruce, and needle-leaved junipers, have buds at the base of each leaf, but tend to bud towards the end of the most recent growth. Last year we dug up a Christmas tree from our allotment and I pruned the ends of most of the branches because it was too wide to fit into the house. The effect of this has been to stimulate the subordinate branches to bud – but again this has only happened at the ends of the branches (see below). Something about firs & spruces keeps the active budding zone at the end of branches.
As well as understanding the budding pattern, a key question for bonsai afficionados is whether or not a particular tree will backbud. That is, will it be possible to increase ramification and foliage density by encouraging axillary or adventitious buds to form.
Gymnosperms were traditionally believed not to resprout, with research in the past finding that buds are not present in leaf axils of conifers. Despite that, there are quite a few gymnosperms species which do, including the following. Some of these “do not have distinct buds at all; they produce new growth from meristematic tissue hidden under the skin of the twig” (Thomas, 2018) – this is known as an epicormic bud. This may be a false distinction since the meristematic tissue may just be early buds which are not developed enough to be visible.
Some Abies (fir)ref including Abies nordmannianaref
Araucaria & Agatha species including including Hoop Pineref and Wollemi pine ref1, ref2
Some Pinus (pines)ref – but pines are notorious for losing their ability to bud anywhere other than on the most recent 1-2 years old stems. Brent Walston at Evergreen Gardenworks says with Pinus thunbergii that as long as there is still a living needle on a stem, if you cut the stem above it, that will force a bud at the needle axil.ref This lines up with the idea that buds in pines are present under the leaf axil of long shoot leaves.
Thujopsis dolabrata (a Japanese species similar to Thuja)
So actually there are quite a few!
Some studies have indicated that “cytokinin sprays on conifers growing in the field can increase the number of visible axillary buds“ref and as a result this study concludes that “conifer leaf axils might not be as blank or empty, at least in recently initiated shoots, as they might appear to be. Cells in the leaf axils, while not forming buds, can maintain a meristematic potential and if they lose meristematic appearance, they may be preferentially able to dedifferentiate into bud forming structures.”ref
In ginkosref, axillary buds are present in the nodes of long shoots only, and these trees can backbud – below is an example of a ginkgo at the Seattle Japanese Garden – you can see new leaves sprouting from the bark of a well-established tree (from the longest long-shoot of all – the trunk).
I’ve also spotted this tree around the corner from my house in London – it was quite tall with all the foliage at the top of the tree – when I saw it cut back so severely I was sure it would die. There were only the tiniest of shoots here are there on the trunk. But in a matter of a few weeks it grew back profusely, which makes me think it must be a Thuja of some kind – perhaps Thuja occidentalis ‘Golden Smaragd’.
Finally another lovely example of conifer resprouting are the amazing dai sugi in Japan – these are Cryptomeria japonica which are cultivated for forestry purposes. The tree is encouraged into a multi-stem form with horizontal branches, which sprout new vertical stems. These are harvested over and over, and new stems grow. In this way the same tree has been used for forestry for hundreds of years without killing the tree. The technique is explained in Jake Hobson’s book Niwaki, which also includes a brilliant photo of bonsai dai sugi, which I think look bizarre but amazing. I have several Cryptomeria japonica at my allotment in the hope of creating something similar (although realistically the ones in this image are probably air-layered).
The term phenology is used to describe the life cycle of a biological organism like a tree. Phenological events for trees include bud development, bud break, flowering, fruiting and leaf & fruit drop, as well as other unseen changes such as sap rising, seed development, root growth, cambial activity or hardening off of tissues for winter.ref
Tree phenology is entwined with the environment in which the tree lives. As there are a very large number of different climates and micro-climates within them, there are accordingly many different nuances in tree phenology, according to the location and environment. Even the same species can show widely different phenology between two different places (at least from a timing point of view).
So to really understand how phenology would play out for your own trees, you need to understand the species phenology and how it varies based on location. You’ll often find bonsai articles are specific to the location of the author which won’t always be relevant to you.
The main phenological events relate to a tree’s growth and reproduction. For example, roots stop growing below 6°C, buds break when the tree detects a low chance of frost in the future (which might damage the tender buds and shoots), photosynthesis, energy production and growth is highest when there is the most sun, and reproduction happens in conditions which most favour seed survival.
In the boreal forests – “high-latitude environments where freezing temperatures occur for 6 to 8 month”ref phenology is mainly driven by temperature, affecting the timing of the start of the growing season and thereby its durationref
Temperate-zone forests are located between the tropics and the boreal forest zone – they have hot summers and cold winters with high temperature variationref, and their phenology is also mainly driven by temperatureref
Mediterranean coniferous forests are mainly driven by water availabilityref
Australian ecosystems are extremely diverse and also subject to irregular events such as fire, drought, cyclones and flooding, which can affect phenological events, but a key driver is water availability.ref Where evergreens dominate in this ecosystem, flowering is the main phenological event.
In tropical forests which have less variation in temperature and usually high water availability, leaf shedding and growth is continuous, but reproduction (flowering and fruiting) demonstrates ‘mast’ timing effects associated with drier than normal conditionsref (ie. all trees fruiting at the same time every seven years)
In boreal and temperate areas the phenology is described in this article and summarised in the images below. But if you’re keen to understand the specific phenology for your tree in your area, you could consult google scholar.
The chart below shows the proportion of Eucalyptus loxophleba flowering at any given time in a seed orchard in the southwest of Western Australia. The highest proportion of flowering happened in spring (Sept-Nov in Australia) but a significant portion also happened in winter (June-Aug). Flowering fell to zero in the hot, dry summer (Dec-Feb).
This all seems a bit confusing given how many different variables there are, but there are some basic principles you can use from a bonsai perspective:
Trees in their growth phase (usually when there is plenty of sun and water) will be able to recover more easily from significant damage (such as large trunk chops or carving wounds) and fight any pathogens which might seek to take advantage of these.
Similarly leaf pruning during active growth will result in more buds activating.
Trees which are in a strong vegetative growth phase (growing leaves and stems) deprioritise root growth. Root growth gets a turn after the leaves establish.
Trees which have set buds but haven’t flowered yet – if you prune indiscriminately – you will lose flowers! There is a way to identify flower buds on your tree but it involves a bit of effort. Flower buds differentiate from vegetative buds at a certain point prior to flowering/leafing out. You can identify different looking buds on your tree, then remove one example of each. Cut it open and look at it under a loupe or microscope and you will be able to see which one was the flower vs the leaf or shoot. Or if you’re both patient and organised, take a picture of some your tree with buds and then with flowers – and you should be able to see what the different bud shapes are.
Storage of carbohydrates to storage tissues will take place during growth phases, and these will be used in turn when less photosynthesis is happening, to drive respiration and other processes requiring energy. Read more about how storage varies in roots here: Root Food Storage (or, can I root prune before bud break?)
If you’re a fan of wiring, doing this before a stem hardens off will allow you more bendability (although watch out for growth around the wire)
Depriving a tree of resources (water, nutrients) will mimic ‘hard times’ and cause it to respond accordingly phenologically – drop its leaves earlier, produce less flowers/fruit or not flower at all, or push out emergency growth (like adventitious buds/suckers)
I think it’s important to say that although the term ‘dormant’ gets used in relation to trees, this is a little misleading. Trees are living organisms and still need to maintain their metabolism even during winter. This includes respiring (using oxygen and stored energy to maintain metabolism), photosynthesising (for any tree with green areas remaining including evergreen trees but also deciduous trees with green stems), transpiring (even deciduous trees still transpire during winter, although a lot less than when they have leaves and in particular they take up water to swell the buds prior to bud breakref), and taking up nutrients through the roots. As I’ve written elsewhere in this site, root growth can happen above 6 degrees C, so your tree may well be more ‘alive’ than you think during winter.
I know there will be people saying at this point – just tell me what happens when!! For those people here are some general guidelines for temperate zones.
You can expect conifers to cease xylem production in autumn and root growth in winter, and to pick these up again between 2-7 degrees C (cambium) and 6-9 degrees C (roots). Buds will burst from early spring onwards depending on the species and latitude and pollen cones will release their pollen. Seed cones will start maturing, which can take just one summer (Picea, Tsuga) or one or more years plus the summer (Pinus, Cedrus). Next year’s buds and future years’ seed cones will form in late summer, and old needles (2+ years depending on species) will drop in late autumn. Mature seed cones will drop or release seed from late autumn onwards. ref1ref2ref3 Hardening leaves for the winter also happens in late autumn.
The main differences for angiosperms in temperate zones revolve around xylem production, leaf growth and senescence within the season, and flowers & fruit. In spring xylem creation will commence – in diffuse porous trees buds can break earlier but ring porous trees need to create the new season’s xylem layer before budding. Some trees will burst bud based on temperature and others on photoperiod (or a combination of the two).ref Whether flowers or leaves come first depends on the species, and the timing of flowers is hugely variable (Frank P Matthews has a list of flowering times for ornamental trees in the UK). The leaves of deciduous trees start a structured senescence process in the autumn, when they remove cholophyll and other molecules from the leaves for storage and recycling (hence the colour changes). After this has been completed the tree creates a cork layer at the base of the leaf causing it to drop off. Fruit develops throughout the growing season and depending on the species will drop off from early summer through to winter.
There’s one more phenological domain which I haven’t covered in this article – the phenology of the microbiome. This is a whole other kettle of…microbes…and might be the subject for a future post.
Finally, the fabulous In ‘Defense of Plants’ podcast has covered phenology in this podcast episode.
After establishing trunk and branch structure, ramification (a fancy word for ‘branching’) of branches and foliage (as well as roots) is a key goal of bonsai. This makes a tree look older and more sophisticated, and gives the bonsai enthusiast options for continued development of the tree.
Ramification is created by branching the stems. Stem branching usually* requires buds, as a new bud creates a new stem. The pattern of stem branching for a particular species will depend on its ‘phyllotaxy’ (leaf morphology) and pattern of buds.
Usually in bonsai we don’t want more than two stems from the same location, the general guidance is to fork into two at any given junction. This is because strong growth of multiple branches at a junction leads to a bulging area on the trunk which bonsai judges don’t like. In the real world, many trees have reverse taper and bulging branch junctions though, so it’s your call. To avoid this situation, remove buds which are in places you don’t want by rubbing or cutting them off.
To improve ramification, you need to encourage as much budding as quickly as possible, then select the buds you want to develop. Pruning the growing tip is the main way to encourage budding, because pruning removes the apical bud (the dominant bud at the end of the stem), diverts resources into buds lower down the stem and sensitises those buds to respond to auxins and develop into shoots. In deciduous trees this should result in at least two buds generating from the stem instead of the one which was there. Another great way to create ramification on deciduous trees is through bud pinching – see Harry Harrington’s detailed explanation of how to do this. Bud pinching removes the entire primary meristem except for two outer leaves, this encourages the buds at those leaf axils to grow, along with two new buds at their bases.
Different species have differing abilities to respond to pruning, so try to get a sense by observing your tree of how well it will cope. Deciduous trees are designed for regeneration so in general they take pruning reasonably well, although if you take it too far they might send out suckers instead of new buds from the branches. With evergreen conifers you want to ensure there is some foliage and at least some buds remaining after you prune, otherwise it may not regenerate (unless it’s a thuja, or a yew, these guys are refoliating machines). I have cedrus seedlings in my collection and by cutting back the apical leader from not long after they germinated, and every year since, they have become extremely bushy and well-ramified (although, at the cost of developing a think trunk).
Gratuitous image of one of my cedar bonsai
Anything which stops or prevents tip extension will drive bud activation and ramification further back on the tree. In the case of conifers, the presence of flowers on the growth tips (as you see in juniper) has this effect as well, and can cause back budding. Lammas growth (a second flush in summer) can give you another round of ramification as long as you’ve pruned beforehand (otherwise it will just add to the existing stems).
Research has found that bud outgrowth is “controlled by plant hormones, including auxin, strigolactones, and cytokinins (CKs); nutrients (sugars, nitrogen, phosphates) and external cues”.ref In particular the sugar sucrose has been identified as a key driver for promoting bud outgrowth and accumulating cytokinins – this is generated by photosynthesis.
In one study on apple trees, foliar application of a synthetically produced cytokinin 6-benzylaminopurine (BA) was found to generate three times the lateral bud growth on currently growing shoots compared to controls (but not on old growth)ref and at the same time reduced the length of the main stem. BA was used to encourage better growth of bean sprouts in China before being bannedref and has been shown to increase the number of leaves (ramification!) on melaleuca alternifolia treesref (melaleuca is the source of tea tree oil), and on some conifersref. Could BA (also known as 6 BAP) be useful in bonsai? You can (like most things) buy this product in foil bags on ebay, but there is a product in the orchid world called Keiki paste which also contains 6 BAP – so maybe some judicious use of ‘crazy keiki cloning paste‘ might also help ramification and shoot development in your trees?ref You can also purchase BAP (as its also known) from vendors involved in hydroponics and suchlike as it’s used in in-vitro plant micropropagation.
If you baulk at paying £18 for 7ml of keiki paste, there is one other source of cytokinins which is a lot cheaper, more sustainable and clearer in its provenance – compost. This study found that compost created particularly from waste collected throughout springref contained 6 BAP. Frustratingly there weren’t any free to read articles analysing compost leachate for cytokinin content, but if it’s in solid compost it’s a fair assumption there are cytokinins in leachate as well. Which makes me feel a lot better about the £300 I recently spent on a Hotbin composter! Which incidentally, produces gallons of leachate, which can be diluted and added as a liquid fertiliser.
* I’ve recently read a study which states that “apical meristems can be surgically divided into at least six parts and these then become autonomous apical meristems.”ref What this suggests is that you could slice growing tips into 6 (or better, two since we don’t want more than two stems from a node) and they would become two stems instead of one! One to try next spring.
** By the way – it’s not auxins which cause apical dominance! Check out page 215 of this book, it’s nutritional status and phyllotaxy which determine the apical stem’s sensitivity to auxin which is present.
This is a rewrite of my original post on shoots, now I know a *lot* more…
So what are shoots? They are the vegetative growth which comes from buds, extending to create new stems. Since stems create the architecture of a tree, shoots are really important when it comes to bonsai.
There are three key concepts to know about when it comes to shoots. The first is the existence of long and short shoots, the second is the way in which different shoots are formed and the third is the concept of the internode.
I had never heard of long and short shoots before researching this site, and I have since found that many articles and books don’t really talk about the fact that many species of tree possess two types of shoots. Shoot differentiation (as it’s known) is present on the vast majority of deciduous angiosperms (flowering trees), all deciduous gymnosperms, and quite a few (around 25%) of evergreen gymnosperms as well, particularly conifers.ref
In these trees, two different types of shoot develop – long shoots and short shoots. Long shoots are exactly as described – they have a terminal bud which continues to build up the length of the shoot over time so it becomes (relatively) long. Short shoots meanwhile don’t persist beyond a limited number of years, they are much shorter than long shoots and have many fewer nodes. Both types of shoots can have leaves, flowers, cones and fruit, but only long shoots can create the long-term architecture of the tree. Importantly, aside from their structural trunk and branches older trees mostly grow short shoots, which is why they look more ramified.ref
In some species (such as pines), short shoots – otherwise known as fascicles – are a feature of the mature vegetative phase of the tree, and don’t appear in the juvenile phase nor with juvenile foliage. An interesting side note is that fascicles can be used to propagate trees with needle leaves, the fascicle is treated like a cutting and placed in rooting hormone and well drained medium – the reason this works is because the fascicle is actually a short shoot and not a leaf.ref
Below is an example of Cedrus libani where the clusters of needles (N1) are on the short shoot (S), and occasionally along the long shoot (L) there are individual needles (N2).
A fascinating – and useful for bonsai – attribute of short shoots is that they almost always have more leaves than the equivalent long shoot. In angiosperms, short shoots have multiple smaller leaves with an almost identical leaf area to a single leaf grown from a long shoot (see example A below).ref And in gymnosperms short shoots have many more leaves and leaf area than long shoots – examples D and E below show the leaves on a short shoot (right hand side) compared to the individual long shoot leaves (left hand side) on larch and dawn redwood.
So hopefully you can see that short shoots are fantastic for ramification! But not so fantastic for building the structure of the tree, since they don’t persist. So how can you tell which is which? Very simply short shoots are smaller, have a lot more leaves, and fall off when their time is up. Often in gymnosperms they will have cones at the end of their leaves.
You may not have realised that whilst the ‘leaflets’ on Cupressaceae species such as dawn redwood, cypress and juniper may appear to be compound leaves, instead they are actually short shoots. When their life comes to an end, the entire short shoot abscises (falls off) along with its leaves. Similarly for pines, what you might know as ‘fascicles’ are actually the short shoots, and on pines only the short shoots bear photosynthesising leaves (needles). Eventually they will fall off.
In angiosperms, a short shoot usually develops from the bud in the leaf axil of the long-shoot leaf, arriving the next season. In gymnosperms, it depends on the species. In Cupressaceae a bud will be sitting at the base of the short shoot so another one should grow once it falls off. In Pinus short shoot buds sit in the long shoot leaves towards the base of the long shoot, and they are positioned at the base of the long shoot bud.ref In Ginkgo both short shoots and long shoots can come from any bud on any type of shoot.
Below is a picture of some interesting behaviour I’d never seen before – this Japanese larch belonging to a member of my bonsai club produced buds and new stems right through the middle of its cones. Pollen and seed cones on larch are terminal organs growing only on short shootsref – which means they aren’t supposed to extend. But Larix is known to be able to change the type of shoot from short to long if damaged (which may have been triggered by the hard pruning it received).ref So in this case what had been a short shoot destined to eventually fall off, instead turned into a long shoot.
So what does it all mean? From a bonsai point of view, the first thing is to work out if a tree has shoot differentiation. If it is deciduous, it will, and if it is a gymnosperm, it still may even if evergreen – gymnosperms which have shoot differentiation include Pseudolarix, Taxodium, Sequoia, Cedrus, Larix, Ginkgo, Pinus & Metasequoia. Understanding the difference between short and long shoots will allow you to understand where foliage will ramify, and where the long-term structure of the tree can come from. On trees which don’t have shoot differentiation, any stem which has a vegetative bud can be used to develop the shape of the tree.
So now we know that long and short shoots exist in many trees, let’s turn to how those shoots form. According to Thomas (2018) , there are three options.
Option 1 is ‘fixed’ or ‘determinate’ growth. These trees preform every part of the shoot in the bud, so they extend very quickly (a few weeks) and then stop. If they are young (less than 10-15 years old) and have the right conditions, they may do this a second time around the start of August (in the Northern hemisphere), this is known as Lammas growth. The shoots from these trees developed based on the conditions at the end of *last year’s* growing season.
Option 2 is ‘free’ or ‘indeterminate’ growth. Thesetrees have only some preformed leaves. Once extended the shoot will continue to produce other leaves from scratch in a continuous fashion. Often these are found in the tropics or warmer climes (my potted Eucalyptus never seem to stop producing leaves even during winter).
Option 3 is ‘rhythmic’ growth. Thesetrees extend in recurrent flushes, with multiple cycles of growth and bud formation during the season.
Outside of the tropics, towards the end of the growing season all trees will stop shoot and leaf growth according to their phenology, in order to complete the formation of buds for next year. If conditions are not good, these buds will be fewer and contain fewer leaves. To see a list of which trees have which types of growth see the Growth Types Table. The relevance to bonsai is that trees with determinate growth are only going to give you one or at most two cracks of the whip in a given season. Those with indeterminate growth might be easier to develop since they will keep extending as long as the conditions are suitable.
Interestingly one study on lammas growth (second flushing) found that 73% of this occurred from lateral buds. We’d obviously love to have this in bonsai as it helps ramification within the same growth season.ref This articleref summarising lammas growth factors says that it can be encouraged by warmer temperatures (Pinus densiflora), extra watering (Pinus sylvestris), nitrogen fertiliser (Pinus sylvestris, Pseudotsuga menziesii) and applying a blackout treatment for less than 2 weeks early in the summer (Picea abies). So from a bonsai perspective see if you can encourage second flushing to generate those lateral buds.
And finally we come to internodes – these are the length of the shoot between each successive leaf. In general bonsai afficionados are looking for short internodes so they can achieve compact, dense foliage. The factors which affect internode length when a tree grows are the same as for any other type of growth – genetics, plant growth regulators and availability of nutrients. Shorter internodes can be achieved by (1) shoot pruning, (2) thigmomorphogenesis and (3) starvation.
If you allow a shoot to extend naturally (and it has no competing stressors), it will prioritise resources into growing as long as it can and the growing tip will suppress the growth of any lateral shoots below it – because the driving force for a tree is to grow large and establish the biggest exposure it can to sunlight. An angiosperm will grow a series of internodes with leaves at each point. What I have observed from looking in my garden is that the internode length on an angiosperm tends to start small (or in some cases leaves are grown directly at the node as well), then increase in size, then reduce again.
To get the smallest internodes, you should prune off the growing tip once the first pair of leaves and the first internode has grown. If leaves have grown at the node, you could remove the shoot altogether (there will be no internode in this case). New shoots will grow from buds in the leaf axils, and if you keep doing this, you will always retain the short first internode and increased ramification.
You could also make use of thigmomorphogenesis which is “the response of plants to mechanically induced flexing, including the brushing or movement of animals against plants, or the flexing of above ground portions of a plant by wind, ice, or snow loading”ref According to this articleref, “the most consistent thigmomorphogenetic effects are a reduction in shoot elongation and an increase in radial growth in response to a flexing stimulus resulting in a plant of shorter stature and thicker, stiffer stem.” i.e shorter internodes and thicker stems.
Thigmomorphogensis is thought to be triggered by plant growth regulators or other substances within the plant signalling when it has been touchedref. To trigger thigmomorphogenesis in your tree, you could expose it to wind while the buds are developing, rub the internodes for 10s daily (seriously, this is what they did in the original studyref which identified the phenomenon), touch the leaves regularly or manhandle the growing shoots.
Another way that bonsai enthusiasts encourage small internodes is by starving the tree. Fertiliser helps the tree grow and this will lead to longer internodes and larger leaves. Holding back fertiliser may result in the desired effect – but also could impact the tree’s health negatively – so it is a balancing act.
So there you have it – shoots turn out to be surprisingly interesting. For your bonsai, try to work out if your tree is shoot differentiated, and if it is, aim to use long shoots for structure and short shoots for foliage ramification. If it has determinate shoot growth, you need to work with the one or two shoot extensions that you get per year, and to get that second flush with lots of lateral buds try using one of the techniques above (warmer temperatures, extra watering, nitrogen fertiliser). Finally keep internodes small with judicious pruning, foliage fondling and holding back fertiliser.