When working with conifers it can get extremely sticky as these trees exude resins from cut stems as well as other organs such as seed cones and needles. We can use our understanding of the chemistry of these resins to work out the best way to dissolve them so we can clean our hands and tools (read on).
Conifer resin helps a tree resist microbial attack, particularly when it is cut, and also acts as a deterrent to herbivory.ref So you can understand why it might need to stick to the stem and cover a wound. Some of the active components of resin which defend against microbes are volatile organic compounds, or VOCs, which evaporate under normal atmospheric conditions.ref This wouldn’t be much use to a plant, so the VOCs are dissolved in non-volatile substances, and resin is this combination of both substances.
One study assessed the composition of resin from 13 species of conifers grown in Taiwan and found that the main non-volatile components were ‘diterpenoids’ – these are organic molecules in the terpene family, shown below. You can also see the volatiles they found in this table – α-Pinene was a common one across species.
https://pubmed.ncbi.nlm.nih.gov/34500678/
To work out how to dissolve such a molecule, we need to know what kind of solvent works against it. The rule is ‘like with like’ – you need a similar molecule to dissolve a substance, specifically as it relates to the electrical charge across that molecule – or its polarity.ref
Water is a great solvent, but only for polar substances – those molecules which have a different electrical charge at one end versus the other.ref Not only are terpenes including the diterpenoids above not polarref, but we already know that water won’t dissolve resin otherwise you wouldn’t be reading this post. For similar reasons soap and water won’t work either, because resin is just too hydrophobic (resisting water).
So we need a non-polar solvent. Unfortunately many of these are nasty substances such as benzene and carbon tetrachloride, which are toxic to varying degrees. They also tend to be produced from crude oil, not exactly a sustainable approach.ref1,ref2
But another non-polar solvent turns out to be plain old vegetable oil.ref
This came to me after remembering my year 12 chemistry teacher explaining how soap works. It stuck in my head that soap is able to dissolve oil because the soap molecule has one end which is attracted to water, and another end which is attracted to oil, which it then disrupts so it can be washed away. So if you can dissolve something in oil first, then you should be able to use soap to wash it away.
And this in fact works really well! Put a decent sized drop of cheap vegetable oil on your hands (you don’t need extra virgin olive oil for this one). Rub the oil thoroughly into the resin and over your hands, and you will quickly see it start to dissolve. Step 2 is to add some hand soap, lather well and rinse. One or two rounds of this will remove even the stickiest, blackest, most persistent of conifer resins. And for tools, you can just use the oil and wipe it off versus washing with soap and water, particularly when you have carbon steel which rusts easily.
Another product which pops up as a recommended one in the bonsai world is this one – SB Invigorator. This product is for pest control and claims to control “Whitefly, Aphid, Spider Mite, Mealybug, Scale and Psyllid.”ref As I have recently added a lot of indoor plants to my collection, these pests are becoming rather annoying, so I have been looking for ways to get rid of them without using toxic chemicals. Would SB Invigorator work?
The main claim for this product is that is uses a “physical mode of action”. However the manufacturer fails to explain what this actually means, so it sort of floats in the ether as a claim without any rationale. A physical mode of action is basically one which physically affects the pests in question. Scraping a pest off a leaf or squashing it with your fingernail would be a physical mode of action. Horticultural oil such as neem also uses a physical mode of action by altering the leaf surface characteristics.ref This method does not rely on poisons, instead it disrupts pests’ ability to move around and/or eat your plants.
What is the physical mode of action in SM Invigorator? Well, there are a couple of clues in the company’s safety data sheet and more in the company’s product manual for commercial users.
The main hazardous component (ie. the one which must be identified on the safety data sheet) is Sodium Lauryl Ether Sulphate (1-3% by volume), also known as SLES. SLES is an ‘ionic surfactant’, basically a detergent and foaming agent. A surfactant is a substance which reduces the surface tension of water of a liquid – on a plant this can make the surface slippery to insects and harder for them to gain purchase on a leaf or stem. In fact plants themselves make surfactants, known as saponins, below is an image of the saponins created by the fruit of Sapindus makorossi in a research study into the subject.ref You can see the foam in the tube, which has been shaken – this is due to the surfactants making it easier for air bubbles to be created.ref
https://www.mdpi.com/2413-4155/3/4/44
Side note – the study identifies a range of plantsref which produce high quantities of saponins, including chickpeas. The saponins in chickpeas result in the sticky liquid left behind when you strain a can of chickpeas – also known as aquafaba. The surfactant properties of aquafaba are used to create meringues and other dishes which require air bubbles, without the need to use eggs.ref
So one of the main ingredients in SB Invigorator is detergent, the likes of which can be found in many consumer detergents. How does this affect pests? According to their product manual, which is published for commercial users, “two separate modes of action have been observed: (1) adult whitefly have been observed to stick by the wings to any surface they make contact with and aphids, juvenile whitefly and spider mite if directly hit are trapped by its wetness. (2) On mealybug an initial application removed the protective wax and a second application controlled them.”
This is why they also promote one of the features of the product being “plant wash for a cleaner, shiny appearance”!
I was interested that the biological control company ‘Dovebugs‘ had contributed to the product safety data sheet. I thought perhaps there were microbes in the product as well. But instead I believe they must have been consulted about the effect of SB Invigorator on beneficial microbes. The company’s informationref states “Studies so far have shown SB PLANT INVIGORATOR to be compatible within an integrated pest management programme where beneficial insects are used.”
On other websites selling this product there are several additional claims which are not listed on the company’s website including:
“SB Plant Invigorator contains naturally elements, such as seaweed”ref [this would act as a fertiliser, particularly good at providing micronutrients]
“improves plant health due to the inclusion of chelated iron and nitrogen fertilisers.”ref [more standard fertiliser]
“Active ingredient: Carbonic acid diamide/urea”ref [source of nitrogen = fertiliser]
“based on a blend of natural ingredients, including surfactants, amino acids, and plant extracts.”ref [as above]
“is a foliar feed that can be used on an extensive range of ornamental and edible plants. The spray contains a wide range of nutrients and micro nutrients that encourage growth and improve the condition and health of the plants when sprayed on the leaves.”ref [foliar fertiliser]
“Consisting of blends of surfactants and nutrients or fatty acids and algae extracts”ref
So if the above are true, in addition to the detergent component, SB Invigorator may also contain liquid seaweed and some fertiliser. Since the product is sprayed on the leaves, it could be acting a a foliar feed (see my article on the effectiveness of these here) as well as a general fertiliser since any runoff would end up in the soil.
On Amazon 500ml of this product is currently £13.45. Assuming their product data sheet reflects the diluted product, with 1-3% of SLES, it’s pretty similar to my eCover washing detergent (with 5-15% surfactants undiluted) which is worth 70p for an equivalent concentration and volume. Let’s say it also has 10% or 50ml of liquid seaweed – based on my Shropshire seaweed purchase recently this would be worth 67p – or to be generous 100ml, which is £1.34. Add to that 50g of Chempak 3 fertiliser (probably way too much since 800g makes 1600L) – worth 63p and you have a grand total of £2.67 for a DIY version.
Now one big caveat here is that the actual proportions of these components may be important, and this company appears to have tested their product – although they have not made their tests publicly available. Since the company is based in Guernsey their financial reports aren’t publicly available either, so it’s not possible to read about their company in much detail. So maybe there is a magic formula which they have perfected and of course there are the costs of management, marketing, packaging, distribution etc.
But, if you can’t afford SB Invigorator, and you wanted to try something similar as a do-it-yourself version, you could do worse than start with the recipe for insect deterrent provided by Jerry Coleby-Williams (a botanist, presenter on Gardening Australia and environmentalist). He says his grandad used to use ‘white oil’ for controlling scale. This recipe suggests mixing half a cup of dishwashing detergent mixed with two cups of sunflower oil, and then using one teaspoon of concentrate mixed into a litre of water. If you wanted to, you could add some seaweed extract and/or fertiliser as well.
Note – I tried a detergent solution to get rid of aphids on some succulents in my indoor plant collection (actually Portulacaria afra) and it made the leaves drop off! I think the solution was nowhere near diluted enough (it was before I read Jerry’s recipe). So do a test leaf before you spray everywhere.
I’ve talked about transpiration in quite a few different posts on this site, but a recent thread on http://bonsainut.com caused me to think maybe I should have a post dedicated to it, so here goes…
Transpiration is the evaporation of water from the leaves of a tree. It’s actually a critical process for trees, because excess transpiration is one of the few ways in which a tree can die; so-called ‘hydraulic failure’ has been identified as the most prominent cause of tree death.ref Hydraulic failure – the failure to access enough water to replace water lost mainly through evaporation – causes cell death, xylem failure and a fatal reduction in photosynthesis. So it’s really important for bonsai practitioners to understand this process.
The main driver of transpiration is not – as you might think – to cool the leaves (although this is one reason for it). In fact transpiration is a by-product, or ‘cost’, of photosynthesis, and it happens because of the way that leaves obtain carbon dioxide. You may already know that plants have small pores called ‘stomata’ which open up to let air – and CO2 – inside the leaf. But you might not have known that gaseous CO2 in air needs to be dissolved in water before it can be accessed by chloroplasts and used for photosynthesis (this is explained in Vogel (Chapter 5 – ‘Leaking Water’). This means that water needs to be available on the surfaces inside the leaf – which means that when stomata open up, this water is subject to evaporation.
Vogel says that “only if the relative humidity is 100 percent will water not be lost…[and]…if the leaf’s temperature is above that of the surrounding air, then water can be lost even at that humidity.” He also says that for every gram of CO2 used by a leaf for photosynthesis, it’s estimated that 125 grams of water is lost.
Smith et al (Chapter 4.10 Movement of Water & Minerals) explain that this evaporation causes a constant flow of water known as the ‘transpiration stream’. As water evaporates from the leaf cells, pressure in those cells is reduced, and this negative pressure causes water from the xylem to move into the space, due to strong mutual attraction between water molecules. That in turn pulls more water behind it and so on. This hydraulic mechanism is responsible for pulling water all the way up the tree from the roots. Actually this process is fundamental to the health of the tree, maintaining cell turgor (stiffness), transporting nutrients, metabolites & growth substances synthesised in the roots throughout the tree, and providing a source of water for the phloem stream which flows in the opposite direction providing energy to the tree. When there is enough water available, all of this works perfectly – when there isn’t, problems arise.
The extent of evaporation from the leaves of a tree is determined by several different factors, which can be divided into environmental, tree-specific physical factors and tree-specific response factors.
The main environmental factor which drives transpiration is the ‘vapour pressure deficit’ (“VPD”) – this is the “difference between the amount of moisture in the air and how much moisture the air could potentially hold when it’s saturated.”ref VPD is a function of both heat and humidity, and provides a measure of how powerful the evaporative force of the air is with any combination of these.ref
Occasionally while writing articles for this blog, I end up in the world of cannabis cultivation. Maybe because they are very motivated to keep their crops vigorous, cannabis growers and their equipment suppliers sometimes have the best data and charts out there! This is just such an occasion, see below for an excellent chart from ‘Ceres Greenhouse Solutions’ showing the VPD for a given temperature and humidity (you can download a copy here). The VPD is low in the blue section and high in the red.
What you will notice is that the relationship between humidity and temperature isn’t exactly linear. Also, VPD increases with higher temperature and lower humidity. Since a higher vapour pressure deficit means there is more ‘pull’ on the water in leaves, increasing temperature and decreasing humidity both increase transpiration – and they reinforce each other, so dry and hot is a high transpiration combination.
Another environmental factor is wind. One study found that wind actually improves water use efficiency, because whilst it does increase transpiration, it also increases CO2 uptake, and the net effect is greater water use efficiency and not less.ref But for the purpose of this article, wind does increase transpiration.
Coming onto tree-specific physical factors – these are all the attributes that relate to the size, shape, position and structure of the tree. In general the more foliage a tree has, the more it will transpire – so a large broadleaf tree will transpire significant amounts on a hot day – in one study they found a large canopy tree in the tropics (Eperua purpurea) transpired up to 1180 litres per day!ref By comparison in the same study, smaller (presumably more shaded) trees transpired a lot less. Thomas (Chapter 2: Leaves the food producers) gives the following figures: “<100L/day in conifers, 20-400 L/day in eucalypts and temperate trees such as oaks, reaching perhaps 500 L/day in a well-watered palm and as high as 1200 litres per day in specimens of Eperua purpurea growing out of the top of the Amazonian rainforest canopy.”
The chart below shows the daily transpiration rate during the growing season for a sessile oak tree in Turkey which measured 18.5m x 34.5m – this maxed out at 160 kg/day (effectively 160L).
As well as the volume of foliage, trees have different stomatal size and density (number of stomata in a given area) which are determined by genetics as well as environmental factors (such as intensity of light and VPD to which they are exposed when developing).ref1ref2 Low stomatal area (ie. density x size) will result in lower transpiration when compared to a tree with higher stomatal area. These researchers measured stomatal area for 737 plant species across 9 forests and at the lower end of the spectrum conifers such as Cunninghamia lanceolata (0.2%) and Picea koraiensis (0.4%) had 100 times less stomatal area than angiosperms such as Viburnum betulifolium (23.77%) or Quercus serrata (21.74%). You can download all their data here. Basically the more stomatal area which is open to the air, the more transpiration there will be.
Many trees have wax plugs in their stomata which reduce their efficiency, and transpiration at the same time. To copy a piece from my article on needle leaves, wax deposits in Sitka spruce stomata reduce transpiration by two thirds but photosynthesis by only one third.ref One study found that 81% of the species they looked at contained such plugs and that wax plugs are particularly numerous in conifers.ref
Another factor is the level of transpiration via bark. This isn’t due to stomatal opening but simply due to partial permeability of bark to air – also genetically determined and due to the presence of ‘lenticels’ – small channels which allow passage of water and air for the metabolism of living cells in the bark. One study on Pinus halupensis found that “Bark transpiration was estimated to account for 64–78% of total water loss in drought-stressed trees, but only for 6–11% of the irrigated trees.”ref This is because bark transpiration is passive and unmanaged, unlike leaf transpiration which can be somewhat controlled by the tree (see below).
Also relevant for individual trees is their position relative to other trees and the sun. Shade will reduce the temperature at the leaf surface and reduce transpiration, a mass of trees together along with undergrowth may increase humidity, also reducing transpiration. A tree standing alone or above others will be exposed to higher temperatures and lower humidity, thus increasing transpiration. Different areas on a single tree will be exposed to different combinations of these factors as well, so rates of transpiration will differ even from leaf to leaf on a given tree.
The final category of attributes which determine transpiration relates to the trees’ ‘behaviour’. That is, how they react to different environmental conditions. As we all know trees may be sessile but they are also incredibly dynamic and can adjust a wide range of parameters of their own biology. The main issue they need to address in this case is losing too much water, which could lead to death. As a result, they change their physiology to manage evaporation as well as water intake at the start of the transpiration stream.
To manage evaporation, trees adjust their stomata based on water availability, changing their ‘stomatal conductance’ to reduce transpiration if not enough water is available.
They do this in a couple of different ways – ‘passively’ and ‘actively’.ref The passive mechanism is where lower water pressure within leaves causes guard cells around the stomata to lose their stiffness, which has the effect of reducing the stomatal aperture. The active mechanism relies on the tree producing abscisic acid (ABA) – this “triggers efflux of anions and potassium via guard cell plasma membrane ion channels, resulting in decrease of turgor pressure in guard cells and stomatal closure”.ref
A study on Metasequoia glyptostroboides found that in most conditions of water availability the passive mechanism was in play, and it wasn’t until prolonged or severe water stress was experienced that the active ABA-mediated mechanism came into play.ref The article explains that different gymnosperm species use different combinations of these passive and active processes to manage a lack of water availability by reducing transpiration. Angiosperms by contrast use a more sophisticated and more recently evolved version of the active process, mediated by ABA.ref
Thomas says that stomata usually close when it is “too cold or dark for photosynthesis” or when the leaves are in danger of losing too much water and wilting”. The consequence of stomatal closing is an associated reduction in photosynthesis – so when a tree is drought stressed, it won’t be generating energy at the same rate as when it was healthy. A study measuring photosynthesis versus stomatal conductance for Pinus radiata (see in the chart below) found there was a roughly linear relationship, as the stomatal conductance increased, so did photosynthesis.
There are several other ways that trees manage their transpiration – by adjusting their root conductance (ability to draw in water), changing their leaf expansion so that there are fewer/more leaves which are smaller/larger in area, pointing exposed leaves downwards during hot periods of the day, changing the root/shoot ratio to match water source to water use and by operating a daily cycle of metabolism which optimises transpiration (eg. increasing their root hydraulic conductance at night when there is lower evaporation, and ‘filling up’ to deal with higher transpiration during the day).ref So they are very much active participants in responding to and controlled their transpiration rate.
But what does it all mean for bonsai? The first thing is, if your tree has plenty of water availability, transpiration should not become a problem, but you need to remember that up to 95% of water use is evaporationref so trees need a lot more water than you might expect. The best way to avoid issues associated with excess transpiration is to supply your trees with all the water they need. This is achieved by regular and sufficient watering, and by using a medium which has some water retention to avoid drought stress – but is also well-draining. A well-draining medium allows you to water more often without the risk of waterlogging roots or creating conditions for pathogens to take hold.
Also – a tree’s ability to handle water loss varies widely depending on the species – Thomas gives the examples of eucalypts and alder as species which cannot control transpiration effectively, and some oaks as species which can. So each tree in your collection will be different.
But let’s consider all the factors explained above that increase transpiration: high vapour pressure deficit (high temperature and/or low humidity), wind, lots of foliage, high stomatal area, clean (unwaxed) stomata, passive bark & leaf evaporation, a sunny/solitary/high position, and a lack of water availability to the roots which activate stomatal closure.
Some of these are adjustable for bonsai. If it’s going to be a hot, dry, windy day then your trees are going to transpire a lot more than normal and if their roots can’t keep up, you need to improve their environment; newly collected and recently root-pruned trees or trees in particularly small or shallow pots will be most affected. You can help them by providing shade (reducing the temperature), increasing humidity, and moving them out of the wind – and obviously by watering. For a temporary period, on a very hot day, it might even make sense to sit pots in water (do not do this for an extended period).
Transpiration can also be a problem in the winter as trees do continue to transpire, albeit at lower levels, even if they are deciduous. As such, they do need water to be available which means you need to keep an eye on moisture levels in pots. If they get dry, water them. If the medium is frozen, this will lock up water and can have a dehydrating effect so in this case you need to also water, ideally when it’s above freezing. Mulch is suggested to avoid hydraulic failure for trees in the groundref, a similar approach can be used for bonsai in pots, to reduce freezing and make more water available to roots. Even at night it is not the case that transpiration completely stops – typically it is 5% – 15% of daytime rates.ref
Balancing the amount of foliage with the roots when repotting or pruning is another important way to help your trees manage their transpiration rates, so that there is enough root mass to meet transpiration demands. Root pruning in the heat of summer should be avoided unless a comparable foliage reduction takes place. If you’ve gone a bit far with the root pruning, use the approaches above – provide some shade, increase the humidity and maintain a watering regime. This is where the bagging method for collected trees comes from – it reduces transpiration by increasing humidity and can be used for trees struggling to recover from a severe root prune.
Anti-transpirant is a product that some bonsai aficionados use. This does what it says on the tin – it is a “film-forming complex of polyethylenes and polyterpenes that when applied to foliage will reduce the moisture vapor transmission rate”ref The active substance is derived from conifer resins. In reducing transpiration these products also reduces photosynthesis, which is a consideration. I’m personally not a fan of disrupting a plant’s natural processes in this way, and successful use of the product depends on the individual tree and product selected (read more here).
Hopefully you can see from all of the above that transpiration is an extremely important concept to understand as a bonsai geek, but one which can be managed, as long as you are aware of the factors at play. Here’s to helping our trees avoid hydraulic failure!
Flat leaves are found across the conifer families including Taxus, Cephalotaxus, and even in Pinus (check out Pinus krempfii) as well as many of the Araucariaceae and Podocarpaceae families. Commonly known trees with this leaf shape include all varieties of yew, swamp cypress, dawn redwood and coast redwood. Here are some examples:
Taxus baccata (Yew)Wollemia nobilis (Wollemi Pine)Cephalotaxaceae (Plum Yew)
Conifer flat leaves are relatively inefficient from a photosynthesis point of view, since water and photosynthates have to travel further to reach the vascular bundle/s.ref As a result, they have better photosynthetic efficiency in humid, low light environmentsref and are more common where it is wet or tropical. In these areas their greater leaf surface can help them attract the energy they need without drying out due to having more stomatal openings on their surface.
The primarily southern hemisphere family Podocarpaceae is super interesting and not one that I have worked with much from a bonsai perspective, although buddhist pine (Podocarpus macrophyllus) is a species commonly found as an indoor bonsai in the UK. (It is hardy down to just below -10°C so could be an outdoor bonsai as well). Podocarpaceae are interesting from a foliage perspective, as they have evolved a wide range of different leaf shapes which in some cases have become similar to angiosperms and take the forms of flattened leaves with modifications that allow them to grow larger. Below are some examples from which it’s clear that this family has found a workaround for the venation constraints of others in the conifers.
One study mapped the structure of Podocarpaceae leaves and created a cross-sectional image for each sub-family, their results are shown below (apologies for the poor resolution but the original paper wasn’t great to start with). You can see that although most species only have one vascular bundle, they also have various mechanisms to get substances to and from it – including the orange ‘organised accessory transfusion’ cells and ‘pitted thick-wall mesophyll’ cells. Interestingly leaf (f) from the image above – the largest leaf – is also shown below and it’s the only one to have multiple vascular bundles (N and O) which is similar to angiosperm leaves.
Whilst the majority of Podocarpaceae are flat-leaved, quite a few members of Araucariaceae have flat leaves as well. The famous Wollemi Pine or ‘pinosaurus’ Wollemia nobilis has flat leaves, as does the equally well-known monkey puzzle tree Araucaria araucana and various others in the family. These trees don’t tend to be common bonsai subjects as their leaves are quite large and observing the Wollemi pine in my garden, they don’t have much variation in their form so wouldn’t be that easy to style.
Also unusual among conifers is the Phyllocladus genus. Plants in Phyllocladus don’t have true leaves at all – any leaves they develop are non-synthetic and ephemeral – ie. they quickly drop off. Instead Phyllocladus use their stems, which have developed a leaf-like flattened form, to photosynthesise.ref These are called phylloclades – see below for some examples from New Zealand:
I know they look identical to leaves – and they contain all the same components including vascular bundles and photosynthesising palisade and mesophyll cells. The nuances of why they are not leaves but flattened branches instead are based on the way they develop and branch, and the relationships between organs – if you want to read the details refer to this excellent paper. But sometimes they may get lumped in with flat leaved conifers.
Which brings me to Taxus or yew. This is a very common bonsai subject, with its own family, Taxaceae. Yew leaves are famous for two things. Firstly, along with every other part of the yew except for its aril (the red ‘berry’), yew leaves are renowned for being extremely toxic. They contain ‘taxine alkaloids’ of which only a small amount is needed to bring on “dizziness, nausea, vomiting, diffuse abdominal pain, cardiac arrest, respiratory paralysis and death” in all animals including humansref. Secondly, they are one of the few conifers which are known to do just fine in shady positions, although they are also happy in full sun as their leaves adjust to different light levels.ref
Taxus have a unique stomatal feature called ‘papillose’ cellsref which can only be described as ‘pimply’ (see below) – basically the entire surface of the leaf has tiny protrusions as part of the cuticle structure. The presence of these pimples is one of the ways of identifying a Taxus leaf. I couldn’t find any clear explanation as to the role of these papillae, except perhaps to provide some level of water-repellence.ref Taxus are known to harbour endophytes (microbes) in their leaves which help combat pathogensref so it could be that the nooks and crannies created by papillae are a nice home for endophytes.
Anyway, what of all this is relevant for bonsai? It sort of depends on which flat-leaved species that you have. Many will be suited to humid and/or low-light environments, so keep an eye on your watering and don’t let them dry out – they will probably appreciate a misting every now and then. Full sun may not be necessary – or may even be over the top depending on your location – but as always find out what a given species needs as there are lots of quirky members of this club with their own unique requirements.
I’ll be honest, I find conifer needles to be quite weird. As someone raised in the southern hemisphere, true pine trees were something we had at Christmas and that was about it (this is true because the most common Christmas tree found in Australia is Pinus radiata). But actually, needles really are just plain old leaves. They contain all the same components as ‘normal’ leaves, like an epidermis, photosynthesising mesophyll cells, green chlorophyll pigment, xylem/phloem, stomata and so on. They just happen to have pushed the ‘leaf’ form to the extreme, ending up extremely long, thin and tough.
Within conifers, what are *called* needle leaves are found across most of the families, including Araucariaceae (Norfolk Island pine, Cook pine, Hoop pine), Cupressaceae (juniper, Thuja, Cryptomeria japonica), Pinaceae (pines) and Podocarpaceae (Platycladus).
At this point though I think it’s important to point out that the Pinaceae family is the least similar to all the other conifer families, having diverged from them very early in evolutionary history. Look at the ‘family tree’ of conifers below and you can see that Pinaceae including pine, cedars, larch, fir and spruce, has been evolving separately for the longest of any conifer family. This means that ‘needle leaves’ in Pinaceae are not the same thing at all as needle leaves in other families, and they probably need to be treated as two separate sub-categories.
This is why it can be confusing to understand what people are talking about when they refer to needle leaves in conifers. Needle leaves in Pinaceae are obvious – they are long, thin, spiky, tough, 3-dimensional and in mature foliage form in clusters known as fascicles which are actually short shoots (see my post on shoots for more on these). What you and I would call pine/fir/spruce needles – as per these examples:
PineCedarSpruce
Needle leaves in other families are a bit more ambiguous. Sometimes they are referred to as ‘awl-shaped’ or ‘sabre-shaped’ and often they have a needly element but also a scale-leafy element. Needle leaves in these families, such as Juniperus and Cupressus, often mature into scale leaves. Here are a few examples of non-Pinaceae needle leaves:
As is obvious, these are different to Pinaceae needles and it’s not just the leaf shape and configuration – one major difference is that these species usually have a photosynthetic stem which pines definitely do not have. You can sort of see how these leaves could change to become scale leaves by the way they are attached to the stem in alternating pairs – if they just shrink and get closer to the stem you could see a mature scale leaf emerge.
For the sake of the rest of this post, I’m going to focus on Pinaceae needle leaves, since these are persistent needles whereas most of the needle leaves on other families are juvenile (although there are some exceptions).
One of the key attributes of Pinaceae needle leaves is that they have a 3-dimensional profile – usually with a quadrangular, triangular or semicircular cross-section, as shown in the images below (1,2,3 & 5 needle pines):
https://www.flickr.com/photos/146824358@N03
Anatomically, Pinaceae needles are like other conifer leaves in only having one or two vascular bundles for water & sugar transport. Their stomata are arranged more or less evenly around the needle and appear in lines. But needles are unique in having an unusual type of mesophyll cell (a cell used for photosynthesis). Instead of cylindrical palisade cells lined up under the epidermis like other plants, needles have frilly looking ‘arm palisade parenchyma’ (see below). These have very lignified (woody) cell walls which intrude into the cells and it is this feature which is thought to provide needles with their extreme cold resistance.
arm palisade cells (pine needle)Pine needle with arm palisade cells
Pinaceae needles are actually *the* most frost-hardy leaves of all. Mountain pine needles survive temperatures down to −93°C and can still perform gas exchange (oxygen for cell respiration) even when their needles are frozen.ref They are also highly resistant to herbivory, due to being tough, spiky, full of toxic resins and not very nutritious. And they live anywhere from 2 to 45 years (Bristlecone pine has the record).ref The needle is one tough leaf!
Pine foliage is also heteroblastic – which means it has one type of foliage during its juvenile phase, which lasts 1-3 years, and a different type during its mature phase. Juvenile needles don’t appear on fascicles, instead developing directly on the stem, and their profile is more flattened – although it’s not super noticeable until you know to look.ref Mature foliage develops in fascicles (bundles) and is the familiar three dimensional profile – they fall off as a group when the fascicle falls off. See examples below – Pinus cembroides juvenile on the left and adult on the right of both frames.
Juvenile and mature needles have different properties with juveniles having 3x the photosynthetic capability of mature needles. This is believed to be able to provide such species with a ‘fast return’ strategy whilst establishing and a ‘slow return stress-resistant’ strategy when older.ref
Surprisingly needle leaves actually have better photosynthetic performance than other conifer leaves – despite that fact that Pinaceae have lost key genes required for photosynthesis in other plants.ref Their improved performance is believed to be because water has to travel further than in non-needle leaves, but there is likely also a genetic factor which hasn’t been discovered yet.ref Needle leaves take a couple of years to reach their full photosynthetic capacity, but once they do, from then on it reduces with age. One study found that for spruce leaves this declines linearly from the 2 year point, reducing to 30% or less of the maximum photosynthetic capacity by the 6 year point.ref
When photosynthesising, trees need to deal with transpiration, which is the evaporation of water from the stomata in their leaves and is the main driver of their water requirements. Needle-leaved trees have a massive advantage in this domain, as many species have wax deposits in their stomata which reduce transpiration. It has been found that wax deposits in Sitka spruce stomata reduce transpiration by two thirds but photosynthesis by only one third.ref Unfortunately these waxes are degraded by pollution, so needle leaved trees can dry out in high pollution areas.ref
Since needle leaves can generate more energy from the light they have, and from a wider range of sun angles, they can survive in poorer light conditions than flattened-leaved species. Along with their cold resistance and ability to minimise transpiration, this is why the boreal forest comprises mainly Pinus, Abies and Picea species, which all have needle leaves.ref
Finally a note on buds and how needle leaves develop on a tree. Leaf buds on mature needle-leaved species form as part of a shoot rather than individually, and these are usually determinate, which means that everything is formed inside the bud. So before bud break the leaf primordia (baby needles) are sitting inside the bud. Below are some cross-sections of long shoot pine buds (which you might know as candles) and a spruce bud (on the right). The brackets show where short shoot buds are located, and within these the baby leaves are waiting to emerge with the shoot.
Contrary to some advice, needles do all their extension in their first year of growth. After this they replace or add to their phloem annually, but not their xylem.ref This means there may be some thickening of needles, but no lengthening after the first year. They may be shorter with increased dryness and poverty of the soil.ref Eventually needles fall off along with their fascicle and the other needles in their group. However, pines have been shown to retain their needles up to twice as long if they have been defoliated (eg. by insects).ref
What’s the impact of this all for the bonsai enthusiast? Firstly your needle-leaved species are going to be tough, they will cope with reduced water, poor soil, wind, rain and freezing temperatures. They definitely do not want to be inside.
Secondly as per conifer leaves in general, needle leaves are not as plastic or regenerative as angiosperms – they are part of a shoot and form in the bud, so there aren’t as many styling options as you find with angiosperms or even flat or scale-leaved conifers. And they have a more relaxed time frame than angiosperms do – needles may stick around for a long time – usually this will be 2-4 years in most species and low-medium elevations but can be a lot higher. So your styling decisions can’t be completely redone on an annual basis – a better approach for needle-leaves is a gradual evolution towards a vision. These trees suit bonsai practitioners with patience and a slow, thoughtful approach.
Bonsai folk like small leaves and in the case of needles, short needles. This is achieved in one of two ways. The first way is that long shoots (candles) are completely broken off early in the growing season. This forces the tree to activate dormant foliage buds at the base of the shoot, which don’t have the time or resources to develop full length needles. I’m not sure whether breaking the part of the candle off (as is also advocated) would also reduce needle size since this retains the active short shoots at the base of the candle to still develop. This practice might have a slightly different effect of creating more short shoots with more needles, so giving denser foliage, rather than shorter needles. The second way to reduce needle size is to starve the tree of water and nutrients, but I’d say manipulating candles would be better for the health of the tree. Obviously to add branches to a tree you need to leave the long shoots in place to develop, as these are what create the long-term framework.
To finish off, I just have to share one of the brilliant images created by Gerhard Vicek who does microscope cross-sections of plants – below is a cross-section of a Cedrus atlantica needle. The beautiful staining he does of his samples makes the different cell types really clear. There are epidermal cells (in red) on the outside, below these the unusual frilly arm palisade cells (in green). Then you have the transfusion cells in a brown & white ring, which move water to the outside of the needle and sugars to the centre. In the centre you can see two vascular bundles with the tiny xylem & phloem cells. Truly his images are art for the bonsai science nerd!
Please visit Gerhard Vicek’s’s website for more great microscope images of trees: foto-vision.at
Scale leaves are a curious form of conifer leaves which cover up the stem in interlocking patterns. I believe they are called ‘scale’ because they look a bit like fish scales in the way they overlap but I have not found an authoritative source which confirms this. Scale leaves appear primarily in the Cupressaceae family – including junipers, various cypresses, Arborvitae/Thuja, redwoods & Callitris, as well as in the Podocarpaceae family including Dacrydium and Acmopyle. Sometimes the scale leaf form is the mature foliage, while the juvenile foliage takes a needle form (see my post on conifer needle leaves).
Some examples of scale foliage leaflets are below:
Juniper with strobiliCypressThuja occidentalis
You’ll note that I called them ‘leaflets’. The actual leaves are the individual scales that you see in the images, which all combine to create a larger leaflet which is actually a short shoot. The leaves are wrapped around and connected to the stem of the shoot underneath.
Scale leaves are usually in opposite pairs, and depending on the species can have main or ‘facial’ scales and lateral scales with slightly different anatomy. Below is a scanning electron microscope image of Thuja occidentalis leaves which demonstrates these two scale types.
Probably the most distinctive attribute of scale-leaved species is the leaf pattern. Each species has a distinct cross-sectional profile, with different leaf shapes and configurations, these are what ultimately create the three-dimensional shape of the leaflet.
Great work was done on this by some Iranian researchers, who created the following cross-section drawings which I have matched to images of the species in each drawing. These show how the scale leaves attach to each stem, the positions of vascular bundles, resin ducts (large holes) and stomata (which are mostly present in the grooves indicated by ‘S’).
As can be seen in the drawings, scale leaves are very simple, usually with a single vascular bundle (other than Juniperus excelsa above which has none), palisade and spongy mesophyll cells for photosynthesis and the darker transfusion cells which move water to and from the stem.ref
In terms of their performance, there actually isn’t a lot of information out there comparing scale leaves with needle or flat leaves. One study found that Thuja leaves were about on par with pine needles in terms of photosynthesisref and another found that juvenile needle leaves of Juniperus sabina outperformed its scale leavesref. Many studies seem to conflate needle and scale conifer leaves and talk about both of them having strong performance in high sun, low humidity situations. There must be some benefit, because quite a few species ‘graduate’ to scale foliage as they age, but I haven’t found any research explaining what that benefit may be.
One weird and wonderful variation of the scale leaf is the unusual ‘axial’ leaf of the coast redwood. Most people associate the coast redwood (Sequoia sempervirens) with a flat-leaved leaflet as shown on the left – and in fact this type of leaf makes up 95% of the leaf surface area of these trees. But 5% is made up of the axial-leaved ‘twiglet’ on the right. These leaves are optimised to absorb water, having much less waxy coating than the flat leaves and contribute up to 30% of the water requirement of the tree. Which can be high, given the size of a coast redwood!
Since scale leaves are associated with older trees, they are preferred for bonsai. If your tree is still in its juvenile phase, you need to let it grow as it’s believed that the trigger for changing phase to mature foliage is the number of meristem cell divisions.ref Pruning the leader on these trees will keep them permanently in a juvenile state, so let the tree grow until it develops mature foliage, then you will need to use all branches & foliage *after* this point to style your tree.
Also remember that scale leaves usually appear on short shoots, which abscise (fall off) as a unit. But don’t worry because usually there will be a bud waiting at the base of the short shoot to replace the one which fell.
Finally if you look at how scale leaves connect to the stem, I believe that the technique of pinching leaves (and stem) off instead of pruning with secateurs or scissors would leave less dead material on the tree. Cutting straight through a stem is always going to sever one or more scale leaves and cause them to die and go brown. The pinching technique is when you use your fingertips, and pull the stem gently so it breaks at a natural breaking point between leaves.
You may have noticed that certain conifer species’ leaves go a different colour over the winter without dying. This phenomenon is most associated with Buxus (box) and Cryptomeria japonica (Japanese cedar) which take on a range of bronze, reddy-brown colours, as shown in the examples below:
Cryptomeria japonica spiralis (from my garden, with frost)Cryptomeria japonica (https://www.flickr.com/photos/oregonstateuniversity/46192395845)Cryptomeria japonica
Although it may look a bit alarming, these leaves will go green again as the temperatures warm up. The reason for the colour change is that in these species, sun-exposed leaves at low temperatures shift their pigmentation to protect themselves from excess energy.
During the winter, evergreen trees face the challenge that the sun can be relatively strong, but the temperatures low. Many of the biological processes needed to convert light into sugars rely on enzymes, which don’t work efficiently at very low temperatures. Although it’s possible for plants to acclimate to lower temperaturesref, beyond a certain point the rate of photosynthesis starts to outpace the downstream processes, and excess energy and reactive oxygen species (“ROS”) accumulate. This can permanently damage Photosystem II (part of the photosynthetic apparatus in plants), so a mechanism is needed to avoid that.
The solution adopted by Cryptomeria japonica is to transform some of its chloroplasts into chromoplasts.ref
The part of the plant cell which photosynthesises is the chloroplast, which is a self-contained organelle with its own DNA known as a ‘plastid’ (and also, in conifers, paternally inherited). Normally it synthesises the green pigment chlorophyll. But plastids can also be ‘chromoplasts’, instead of synthesising chlorophyll, they synthesise coloured carotenoidsref which aside from being different colours, also act as anti-oxidants, removing dangerous excess oxygen levels from plant tissues. The pigments found in winter Cryptomeria foliage include Rhodoxanthin (found in Taxus baccata arils), Zeaxanthin (found in marigolds) and Antheraxanthin (found in dandelions).ref These are all carotenoids which are known to safely dissipate excess energy during photosynthesis.ref
Red Taxus arils coloured by Rhodoxanthin (image: Kew Gardens)Marigolds coloured by Zeaxanthin (image: Wikipedia)A dandelion coloured by Antheraxanthin (image: Wikipedia)
So during winter when temperatures fall, green chloroplasts are converted into coloured chromoplastsref, and vice versaref when the temperature rises. The effect of this temporary transformation is to reduce photosynthesis and to increase the plant’s anti-oxidant levels, so that excess energy is dissipated safely and the photosynthetic apparatus is not damaged.
Aside from being observed in Cryptomeriarefand Buxusref, there is also anecdotal evidence that the same process happens in Juniperusref and Thuja (see below for one of my Thuja bonsai – the white bits are frost and the foliage is a distinctly reddy-brown colour). Interestingly these are all ‘flattened’ forms of conifer leaves, and colour-changing behaviour is rarely observed in needle-type leaves, which may instead deal with the problem of winter sun by plugging up their stomata to reduce the rate of photosynthesis. One exception is Pinus contorta ‘Chief Joseph’ which has golden needles in the winter and green ones in the summer.
The conversion of chloroplasts to chromoplasts is also responsible for the colouring of fruit and flowers, but it’s *not* what makes leaves coloured in the autumn (that would be ‘gerontoplasts’ref).
It seems that the conifer species which can do this are all members of Cupressaceae. So if you see pine needles going brown, that is probably the needles dying (which they do naturally after a certain period of time) unless it is a rare form. But if your Cryptomeria, Juniperus or Thuja develops a winter ‘blush’, don’t worry, it will probably come back all green when the temperature warms up.
I’ve been planning a post on this subject for a while because conifers have always been a bit scary to me from a bonsai point of view – they don’t seem as forgiving or obvious in terms of their growth behaviour. This was one of those subjects which ended up being a lot more interesting and complex than I was expecting – once I hit 3000 words for this post I realised I needed to separate things out! So below is a *summary* overview of conifer leaves, and detail on the three different types of conifer leaves are in separate posts: conifer needle leaves, conifer scale leavesand conifer flat leaves.
But let’s start from the start. What are conifers? Strictly speaking they are any of the species in the family Pinopsida also known as Pinales or Pinophyta (for a reminder review the previous post on The kingdom Plantae and where trees fit in), that is to say, the Pinophytes.Pinophytes are cone-bearing plants, hence the name conifers. They include six different families:
Araucariaceae (including monkey puzzles and the Wollemi pine)
Podocarpaceae (mainly southern hemisphere evergreens including Buddhist Pine), including Phyllocladaceae (celery pines from New Zealand)
Sciadopityaceae (Japanese umbrella pine is the only member in this family)
Taxaceae (yews) including Cephalotaxaceae (Japanese plum yew)
So why do these families have different leaves to those of angiosperms/flowering plants? It’s because gymnosperms (including conifers) and angiosperms diverged in their evolutionary paths 350 million years agoref and as a result they have evolved with key genetic differences. These are exposed in leaves in five key areas:
Venation – the structure of the vascular system which transports water through the leaf and products of photosynthesis back into the tree (ie. its ‘veins’) (and thus determines the possible leaf shapes)
Stomata– the distribution, density and effectiveness of the pores on the leaf which allow air in and water vapour/oxygen out
The photosynthetic apparatus – how the cells in the leaf are arranged to perform photosynthesis and which reactions are used
Heteroblasty – the phenomenon of ‘extreme variation in leaf morphology during plant development’ or in other words, leaves being completely different on young plants versus old plants of the same species (trees which have different juvenile and mature foliage) – although this also exists in angiosperms the versions in conifers are unique genetically
Resin canals – the ducts in conifer leaves & stems containing secondary metabolites
Starting with venation, the vascular system of conifers (which performs water & sugar transport) has only one single vein or two parallel veins per leaf, running up its centre. This is shown in the examples of conifer leaf cross-sections below – purple shows the xylem (water transport) and the blue shows the phloem (sugar sap transport). (2), (10) and (14) have two parallel sets of veins and (5) & (12) have a single, larger vein.
By contrast the vascular system in the leaf of a flowering plant is much more sophisticated with many different vein patterns across speciesref and the average vein length per area in an angiosperm leaf is 2 to 5 times higher than in conifer leaves.ref Some examples of angiosperm leaf venation are below – you can see veins branching and extending to every part of the leaf and this is one of the advantages that allow angiosperms to create larger leaves (hence the name ‘broadleaf’).
The vascular structure of conifer leaves limits how much water can be delivered to their outer edges. From the vascular bundle/s, ‘transfusion tissues’ or specialised cells conduct water and photosynthates to and from the margins.ref Their conducting capacity is limited, which in turn limits how wide a leaf can become. In layman’s terms, because conifer leaves have basic water piping, they can’t grow too wide – which affects the size and shape that conifer leaves can take.
Conifer leaf shape categorisation is inconsistent across the literature, and you may see different descriptions such as awl-shaped, sabre-shaped or even intermediate (a catch-all for anything which doesn’t fit). A reasonable set of descriptions has been created by Paul Fantz at the North Carolina State University. But at the end of the day most conifer leaves fit into one of three types – flat, scale or needle leaves. A nice study was done in Iran which produced line drawings of the main three types of conifer leaf, which you can see below (and here). On the top is a flat leaf of Taxus baccata (yew), on the bottom left is a scale leaf (and stem) of Cupressus sempervirens (italian yew) and on the bottom right is a needle leaf of Juniperus communis (common juniper). Due to their shapes, each type of leaf is a little bit different in terms of how they perform in a given environment, and you can learn more about this in my posts about each type: flat, needle, scale. The fact that the same tree produces foliage of more than one type is covered below in the section about heteroblasty.
Now let’s consider the stomata on conifer leaves (to learn or remind yourself about stomata you can read my stomata post). Whilst conifers have the same basic structure for their stomata, with one guard cell on either side, they differ from angiosperms in their arrangement and effectiveness.
Conifer stomata develop at the base of each leaf, meaning that they grow out in longitudinal bands as the leaf emerges, whereas angiosperm stomata develop at multiple points on a leaf, resulting in more variation in their patternsref. In needle species they are arranged around all sides of the leaf (with a few exceptions), in scale leaf species they appear in the grooves between scales and stem, and in flattened leaf species they appear mainly on the bottom of the leaf. Below is an image of the stomata from a Picea species, showing them arranged in lines:
Stomata in conifers have a couple of other characteristics – often they are ‘sunken’ or set into the layers of the leaf, as well as filled with wax plugs.ref This massively reduces the gas exchange capacity of the leaves – one study found that gas exchange was only 35% compared to species without wax plugs. Their conclusion was although this blocks the stomata and reduces photosynthesis, it may have been an advantage during wetter periods of earth’s history by keeping the pores free of water. The wax plugs also prevent fungal intrusion – which is more of a risk for conifers with long-lived leaves. Finally a less open stomata also reduces water loss. This allows conifers to survive in drier areas and to stay alive for longer with minimal water – hence they are now found in more extreme environments where angiosperms can’t survive. Below is a sunken stomata from a Tsuga canadiensis on the left and a Cryptomeria japonica stomata full of wax on the right.
Next we need to look at one of the most important attributes of a leaf – its photosynthetic apparatus and performance.
Whilst conifer leaves photosynthesise about 30% less effectively than angiosperm leavesref, they live and photosynthesise on average 50% longer when compared to angiosperm evergreens – and obviously much longer (around 300%) when compared to deciduous angiosperm leaves.ref So overall conifers need to invest less resources to generate their energy, since each leaf works for longer periods. Where angiosperm leaves have a ‘live fast, die young’ lifestyle, conifer leaves are more ‘slow and steady wins the race’.
One surprising fact I came across while researching this post was that conifer seedlings can actually grow in the dark. They are able to synthesise chlorophyll and create the photosynthetic apparatus without light, and these are ready to work as soon as the plant is illuminated- although the amount of chlorophyll produced is lower than if the seedling has been illuminated.ref This makes sense since seedlings may often germinate in low light conditions on a forest floor.
Like angiosperms, conifers can have different shade and sun leaves (this is known as ‘heterophylly’). In Abies alba (silver fir) sun leaves are on average longer, have thicker cuticles, more photosynthesising palisade mesophyll cells, fewer spongy mesophyll cells and more stomata than shade leaves, as well as significantly higher photosynthetic performance.ref By contrast shade leaves contain 3 times more chlorophyll content and 2.5 times more carotenoids than sun leaves. Even the arrangement of sun and shade leaves look quite different – see the image below showing sun leaves on the left and the shade leaves on the right.ref
Another factor which determines the photosynthetic performance of a leaf is its age. Except for the few deciduous conifers, conifer leaves can last anywhere from one to 45 years, although the latter is unusually long. The data is scattered across many papers but to provide some examples, the majority of pine needles live for 2-8 yearsref , the scale leaves on Thuja plicata live on average 8 yearsref, and flat yew leaves also live up to 8 years.ref Needle leaves live longer at higher elevations and with poorer conditions in general (such as lack of water).ref
Which brings me to the topic of heteroblasty, or trees which have obviously different juvenile and mature leaves. It’s a well noted phenomenon in bonsai circles that certain junipers have needle leaves when young and scale leaves when older. It turns out that heteroblasty is observed in Cupressaceaeref, Pinaceaeref and Podocarpaceaeref and results from what is called a ‘phase change’ in the shoot apical meristem. This is when the growing tips change to produce different organs – so instead of producing buds that become juvenile leaves, they produce buds which become mature leaves – and eventually buds which become reproductive organs as well. This phase change is relatively stable, so once a meristem produces mature foliage, it will continue to do so. It is also position specific – so the lower branches may retain juvenile foliage even when the rest of the tree has mature foliage.ref
One explanation for heteroblasty is that it’s a useful way for plants to deter herbivores or other environmental hazards that exist for smaller, younger plants. New Zealand has a high number of heteroblastic plants (200 species), and academics have proposed that the unusual branching form in juvenile trees which is specific to the area has specifically developed to deter large ratite birds like emus and moa.ref 10 such species were found which changed their leaves and branches once they surpassed 3m in height (the maximum bite-height of the ratites). However since there are no more moas, it’s hard to prove the theory, which is apparently hotly debated.ref
Phase changes are controlled by genes and plant growth regulators, which change their expression when a meristem has undergone a certain number of cell divisions.ref This was demonstrated by showing that mature flowering meristems, when rooted as cuttings, also flowered and so retained their mature state. This is why position matters when it comes to heteroblasty and only meristems which have reached the mature phase will produce mature foliage.
Since phase change to a juvenile state is desirable for plant cloning, there are studies which have considered how to maintain juvenility or reverse it in mature plants. One method for delaying phase change is to ‘hedge’ – what you and I would call pruning – presumably because this removes the apical meristem programmed for the new phase and reverts to meristems lower down the plant which haven’t changed phase. Another is to apply stress to a plant by starving it, dehydrating it or exposing it to heavy metals.ref
The final and fifth familiar attribute of conifer leaves that differs from angiosperms is that they are almost all resinous. Conifer resins are mostly terpenes made up of linked isoprene elements (C5H8) and are conducted through leaves (as well as some cones and wood) through resin canals. 30,000+ different terpene structures produced by conifers have been identified – some of which are used to produce various products including turpentine, printing inks, soap, plastic, fireworks, and tar. The effect of resinous leaves is to deter insects (Farjon, 2008) and microbes.ref Resin doesn’t feature too much in bonsai (other than when you’re cleaning your branch cutters), but the resin does provide a defensive benefit to your trees which is probably better than many of the chemicals that are sold for the purpose.
Anyway what does it all mean for bonsai? (Thank god I hear you say – it only took her 2000 words!!)
Well let’s start by acknowledging that conifer leaves are quite different from those of angiosperms. Their vascular system dictates that the leaves take one of the three forms – needle, scale or flattened, and aside from the few deciduous conifers, in general their leaves are designed to stay on the tree for much longer than most angiosperms. This means you’re not going to get the same level of leaf turnover on your coniferous bonsai as you would with your angiosperms, and your styling decisions need to be more carefully made and executed. It is going to take longer to fix a mistake on a conifer.
Similarly, their photosynthetic rate is not as high as an angiosperm, so in many cases a conifer is not going to be able to achieve the same growth rates as an angiosperm unless they have a lot of light, although there are some more fast-growing species. As per the previous point, conifers are less forgiving of poor styling decision.
Depending on its leaf type, your different conifers will prefer different conditions (full sun for needle, humid and less sunny for flattened), but you should also be thinking about how to cultivate the types of leaves you want to see on your tree. Sun needles appear denser and better for bonsai, so shading a fir or a pine is probably not a great idea. Similarly making use of short shoots with their increased leaf numbers is important (see my post on shoots).
Species which display needle-scale leaf heteroblasty are a special case as usually you want them to take on mature scale foliage which is preferred in bonsai. To do this, lower, older branches (with the juvenile form) will eventually need to be removed, and you shouldn’t prune the apical stem of these species until they have reached the mature foliage phase. Or sidestep the juvenile phase altogether by taking cuttings of mature foliage which should stay mature unless they are seriously stressed.
A final point would be to say that although conifers all fall under Pinopsida (etc) they have a much longer evolutionary path than angiosperms and more divergence between them, so lumping them all together into one post is not really comparing apples with apples (hehe). So have a look at the other posts which spawned from this one to dive into a bit more detail: conifer needle leaves, conifer scale leaves and conifer flat leaves.
The phyllosphere is the community of microbes which live in and on a plant’s leaves. I had no idea that this even existed before writing the section for this website about the microbiome. Of course, if you think about it for a microsecond, it must! Our world has more microbes than anything else by several orders of magnitude, so, there must be microbes in a tree’s leaves. But the phyllosphere has been less publicised due to the intense interest in the rhizosphere (root microbiome) and in its beneficial microbes which can help plants grow by manipulating the soil and root environment.
The phyllosphere is different to the rhizosphere in that its main microbial members are bacteria and not fungi, although fungi are present, along with some archaea. It has been estimated that there are 1 million -10million bacterial cells per cm2 of leaf surface.ref And worldwide, the phyllosphere is an important microbiome, with a possible 1026 cells! But it’s a relatively hostile environment, with fluctuating temperature & humidity and limited nutrients on the leaf surface. The shape and structure of the leaf at a microscopic level provides a range of microhabitats for bacteria, including the bases of trichomes, stomata, hydathodes (leaf pores), grooves along the veins, epidermal cell junctions, and cuticle depressions.ref A study into tree phyllospheres found 129 bacterial species were significantly associated with the gymnosperms including Armatimonadetes, Actinobacteria, Bacteroidetes, Acidobacteria, TM7, TM6, Deltaproteobacteria, OD1, Fusobacteria, and FBP and 79 with the angiosperms including Chlamydiae, Proteobacteria, Gammaproteobacteria, Alphaproteobacteria, and Firmicutes.ref
Bacteria on a leaf surface, from: https://www.ethlife.ethz.ch/archive_articles/090915_blattleben_kw/index_EN.html
What determines the microbial mass and mix on leaves is a combination of different factors, including the nitrogen content of leaves, the specific leaf area (related to carbon availability), wood density and seed massref and the largest part of the variation seen between phyllospheres comes down to the host species. Conifers have a different phyllobiome than other species, for example they have less ice nuclei active bacteria (bacteria which can cause ice crystals to form) and they have Frankiaceae which is involved in nitrogen fixing in the soil.ref Location also plays a role, with urban trees displaying a different phyllosphere makeup – correlated to ultrafine particulate matter and black carbon on the leaves.ref
Bacteria usually require an available carbon source. You might be surprised to know that similar to roots, leaves also produce exudates (substances they exude into the environment). These include a wide range of carbon compounds, such as carbohydrates, amino acids, organic acids, and sugar alcohols, primarily products of photosynthesis, as well as proteins, oils, secondary metabolites and mucilage.ref These carbon sources are not the only ones – the Methylobacterium species can use methanol exuded from the leaf from the breakdown of pectin as its only carbon source.ref One of the bacterial families found on birch – Rhodospirillaceae – is able to photosynthesise, removing the dependence on leaf carbon sources. Another study discovered that certain phyllosphere bacteria can use diesel for their carbon source!ref
Similarly, bacteria in the rhizosphere produce a range of substances just like they do in the rhizosphere – biosurfactants which reduce surface tension, degrade hydrocarbons and improve moisture levels and dissolved nutrients on the leaf surface, plant growth regulators which open up the leaf cells and cause them to leak nutrients, enzymes which help break down nutrients and protect the bacteria from solar radiation, and phytotoxins (if the bacteria is a pathogen).ref
The benefits of phyllosphere microbes to their host are similar to those in the rhizosphere – for example Acetic Acid Bacteria have been found to perform nitrogen fixation within the needles of Pinus flexilisref, others confer resistance to Bursaphelenchus xylophilus-induced pine wilt diseaseref, some phyllosphere fungi produce zeatin, a cytokinin (plant growth regulator)ref and others auxins, some also produce anti-freeze proteins which lower the freezing temperatureon the leaf.ref Bacteria are implicated in the bioremediation of harmful chemicals or pollutantsref, improved tolerance to stress, production of proteins which trigger the plant to mount defences against pathogens as well as those which attract populations of beneficial fungi.ref
So, just like the rhizosphere, the phyllosphere is a very active place with many microorganisms playing different roles and constantly interacting in a dynamic ecosystem. What this means for bonsai is that there likely are organisms in the foliage which benefit your plant. Similar to the advice in general around the microbiome, applying fungicides, anti-bacterials and chemical pesticides can kill phyllosphere organisms so avoiding this is a good idea.
The reproductive system and organs of plants are extremely varied and complex, and worthy of an entire website to themselves – a comprehensive view is beyond the scope of this website. But what I want to do is provide a bit of information to help you identify which buds might be reproductive vs vegetative.
There are different buds for vegetative (shoots & leaves) and reproductive (flowers, strobili) organs – within each bud a different set of components develop depending on what kind of bud it is. For angiosperms, it’s hypothesised that flower buds are based on the same structures as vegetative buds – that is, a bud starts as vegetative and then differentiates into a flower bud.ref For gymnosperms, reproductive buds contain male or female strobili.ref These are the male pollen cones or the female seed cones.
There are two ways to work out which bud is which – by their appearance or by their position on the tree. The appearance route is best aided by dissecting some actual buds from the tree you are interested in, so you have real data from the real tree. Otherwise, read on for more information about how vegetative and flower buds differ in appearance.
Vegetative buds are “encased by strong, coarse, mature scale leaves. Thinner, more membranous scale leaves make up the next layer. The scale leaves form a protective enclosure surrounding the developing foliage leaves.”ref In many of the articles online, leaf buds are said to be thinner than flower buds (at least for angiosperms). Below are the vegetative buds of Acer pseudoplatanus and Fraxinus excelsiorref:
A flower bud develops sepals, petals, stamen, pistil, ovaries & anthers, instead of leaves and more buds. Below is a scanning electron micrograph of a Bing cherry flower bud forming where M is the meristem, B is the bract and F is the very start of the flower forming – the progression over time is from left to right. On the right is the pistil with ovary (O) style (SY) and stigma (SM). The scale of the image is provided by the white bar on the bottom right hand side, which is 100 micrometres (or microns) – about 1/100th of a centimetre. Obviously it’s impossible to detect a flower bud at this stage by eye, it’s way too small!
The buds of these Bing cherries started forming and were detectable under a microscope from mid-May (the location was Washington state USA) – the year before they would flower and fruit. At a lower magnification below is a progression in flower bud development of a Camellia – the key difference is that at A2 when the flower starts to differentiate, the tip of the bud becomes more rounded and flattens.
Some of the trees we use in bonsai are dioecious which means they are only male or female and not bothref. This means they will produce only one type of flower or strobili bud. To check whether your gymnosperm is dioecious or monoecious you can check the gymnosperm database. Unfortunately I don’t have a reference link for angiosperms.
In gymnosperms there are no flowers, instead the reproductive organs are male or female strobili (pollen or seed cones respectively). A brilliant reference for some of these is available online here. An extract from the publication is provided below with some examples of vegetative and male & female strobili buds.
As noted another way to determine the type of bud is by its position. To do this you need to work out what the architecture of your tree is and where buds of different types typically form. Trees conform to one of 24 architectures as described in Tree Architectural Models and these give an indication to the location of reproductive buds. Trees can have one model for their juvenile phase (before they flower or adopt mature foliage) and then one for their reproductive phase.
In the case of apple, it has been found to conform to Rauh’s model when juvenile and Scarrone’s when reproductive.ref Even with this, “great differences exist between cultivars whether they belong to “spur” vs. “spreading” or “terminal bearing” growth habit”ref
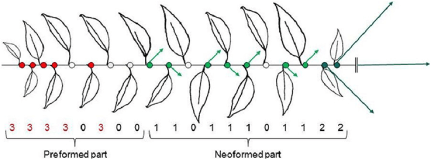
In cherry, “The long branches bear short shoots, also called spurs, in lateral positions on the distal half or two thirds of the branch with more vigorous spurs toward the distal part [the most distant part]. Flowering occurs in axillary positions on the five to six basal-most nodes of all shoots whether long or short. Floral buds are thus located exclusively on the preformed nodes of the previous year shoots.”ref
Red dots are flower buds, formed the previous year, and green dots are vegetative buds, formed during the growing season https://www.frontiersin.org/files/Articles/105157/fpls-05-00666-r2/image_m/fpls-05-00666-g001.jpg
In some species, particularly pines, the long shoot buds have many different components on them in a specific order. Here is a Pinus contorta bud – you can see the male (pollen) cones appear first, and are positioned at the bottom of the final shoot, and the female (seed) cones appear last just below the top of the shoot.
As you can tell from the above, there is quite a bit of complexity in understanding the architectural model of a tree, and where different buds form, as there are many variables involved. Observation is probably going to be the better method.