Another product which pops up as a recommended one in the bonsai world is this one – SB Invigorator. This product is for pest control and claims to control “Whitefly, Aphid, Spider Mite, Mealybug, Scale and Psyllid.”ref As I have recently added a lot of indoor plants to my collection, these pests are becoming rather annoying, so I have been looking for ways to get rid of them without using toxic chemicals. Would SB Invigorator work?
The main claim for this product is that is uses a “physical mode of action”. However the manufacturer fails to explain what this actually means, so it sort of floats in the ether as a claim without any rationale. A physical mode of action is basically one which physically affects the pests in question. Scraping a pest off a leaf or squashing it with your fingernail would be a physical mode of action. Horticultural oil such as neem also uses a physical mode of action by altering the leaf surface characteristics.ref This method does not rely on poisons, instead it disrupts pests’ ability to move around and/or eat your plants.
What is the physical mode of action in SM Invigorator? Well, there are a couple of clues in the company’s safety data sheet and more in the company’s product manual for commercial users.
The main hazardous component (ie. the one which must be identified on the safety data sheet) is Sodium Lauryl Ether Sulphate (1-3% by volume), also known as SLES. SLES is an ‘ionic surfactant’, basically a detergent and foaming agent. A surfactant is a substance which reduces the surface tension of water of a liquid – on a plant this can make the surface slippery to insects and harder for them to gain purchase on a leaf or stem. In fact plants themselves make surfactants, known as saponins, below is an image of the saponins created by the fruit of Sapindus makorossi in a research study into the subject.ref You can see the foam in the tube, which has been shaken – this is due to the surfactants making it easier for air bubbles to be created.ref
https://www.mdpi.com/2413-4155/3/4/44
Side note – the study identifies a range of plantsref which produce high quantities of saponins, including chickpeas. The saponins in chickpeas result in the sticky liquid left behind when you strain a can of chickpeas – also known as aquafaba. The surfactant properties of aquafaba are used to create meringues and other dishes which require air bubbles, without the need to use eggs.ref
So one of the main ingredients in SB Invigorator is detergent, the likes of which can be found in many consumer detergents. How does this affect pests? According to their product manual, which is published for commercial users, “two separate modes of action have been observed: (1) adult whitefly have been observed to stick by the wings to any surface they make contact with and aphids, juvenile whitefly and spider mite if directly hit are trapped by its wetness. (2) On mealybug an initial application removed the protective wax and a second application controlled them.”
This is why they also promote one of the features of the product being “plant wash for a cleaner, shiny appearance”!
I was interested that the biological control company ‘Dovebugs‘ had contributed to the product safety data sheet. I thought perhaps there were microbes in the product as well. But instead I believe they must have been consulted about the effect of SB Invigorator on beneficial microbes. The company’s informationref states “Studies so far have shown SB PLANT INVIGORATOR to be compatible within an integrated pest management programme where beneficial insects are used.”
On other websites selling this product there are several additional claims which are not listed on the company’s website including:
“SB Plant Invigorator contains naturally elements, such as seaweed”ref [this would act as a fertiliser, particularly good at providing micronutrients]
“improves plant health due to the inclusion of chelated iron and nitrogen fertilisers.”ref [more standard fertiliser]
“Active ingredient: Carbonic acid diamide/urea”ref [source of nitrogen = fertiliser]
“based on a blend of natural ingredients, including surfactants, amino acids, and plant extracts.”ref [as above]
“is a foliar feed that can be used on an extensive range of ornamental and edible plants. The spray contains a wide range of nutrients and micro nutrients that encourage growth and improve the condition and health of the plants when sprayed on the leaves.”ref [foliar fertiliser]
“Consisting of blends of surfactants and nutrients or fatty acids and algae extracts”ref
So if the above are true, in addition to the detergent component, SB Invigorator may also contain liquid seaweed and some fertiliser. Since the product is sprayed on the leaves, it could be acting a a foliar feed (see my article on the effectiveness of these here) as well as a general fertiliser since any runoff would end up in the soil.
On Amazon 500ml of this product is currently £13.45. Assuming their product data sheet reflects the diluted product, with 1-3% of SLES, it’s pretty similar to my eCover washing detergent (with 5-15% surfactants undiluted) which is worth 70p for an equivalent concentration and volume. Let’s say it also has 10% or 50ml of liquid seaweed – based on my Shropshire seaweed purchase recently this would be worth 67p – or to be generous 100ml, which is £1.34. Add to that 50g of Chempak 3 fertiliser (probably way too much since 800g makes 1600L) – worth 63p and you have a grand total of £2.67 for a DIY version.
Now one big caveat here is that the actual proportions of these components may be important, and this company appears to have tested their product – although they have not made their tests publicly available. Since the company is based in Guernsey their financial reports aren’t publicly available either, so it’s not possible to read about their company in much detail. So maybe there is a magic formula which they have perfected and of course there are the costs of management, marketing, packaging, distribution etc.
But, if you can’t afford SB Invigorator, and you wanted to try something similar as a do-it-yourself version, you could do worse than start with the recipe for insect deterrent provided by Jerry Coleby-Williams (a botanist, presenter on Gardening Australia and environmentalist). He says his grandad used to use ‘white oil’ for controlling scale. This recipe suggests mixing half a cup of dishwashing detergent mixed with two cups of sunflower oil, and then using one teaspoon of concentrate mixed into a litre of water. If you wanted to, you could add some seaweed extract and/or fertiliser as well.
Note – I tried a detergent solution to get rid of aphids on some succulents in my indoor plant collection (actually Portulacaria afra) and it made the leaves drop off! I think the solution was nowhere near diluted enough (it was before I read Jerry’s recipe). So do a test leaf before you spray everywhere.
I live in London, a city sitting on a giant chalk deposit which formed in the Cretaceous period and stretches all the way to France (via the Eurotunnel)ref Chalk is a form of limestone made up of the shells of marine organisms, and is comprised mostly of Calcium Carbonate (CaCO₃).ref According to my water supplier (Thames Water) “When your drinking water seeps through this rock, it collects traces of minerals like magnesium, calcium and potassium. This is what makes it hard.”ref
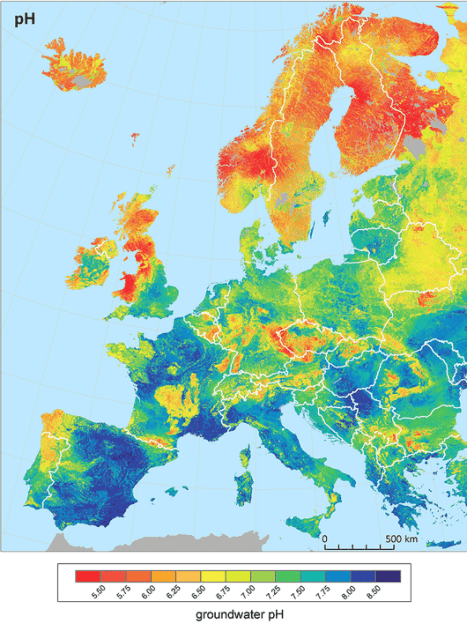
As you can see the water in my area is towards the harder end of hard. But there are plenty of places in Europe with hard water as well, as you can see in this map which comes from a study measuring groundwater in 7,577 sites across the region – most areas in fact are hard with exceptions in Scandinavia, Scotland and northwestern Spain (where igneous/volcanic bedrock dominates)ref:
What is also interesting from this research paper is the corresponding map of groundwater pH (see below). Groundwater pH determines your tap water pH if that’s where your drinking water comes from. Some areas source their drinking water from surface water as well, such as lakes and running watercourses – for example in Sweden it’s 50/50.ref
pH is closely associated with water hardness, with higher levels of calcium carbonate leading to increased pH (in the world of agriculture a common practice to raise the pH of acidic soils is ‘liming’ – or adding calcium carbonate)ref. Look at the areas in Southern Spain and France below which are pH 8 and above – their groundwater is also hard as shown in the map above.
The water in my taps is pH 7.75, so getting close to 8 which is relatively high. Not only that, but continued watering and drying of a bonsai medium with calcium-carbonate-rich water could increase the concentration of calcium carbonate in the pot and potentially make the pH even higher. But is this a bad thing?
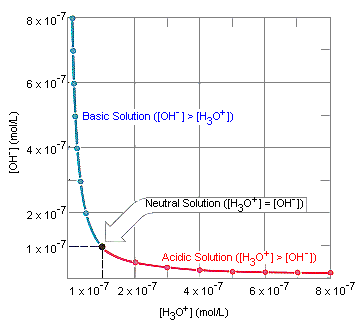
To answer that question we need to take a detour into pH and what it actually means. At this point you can be thankful that usually I wait for a couple of days before posting, because otherwise you’d be deep in the weeds of ions, acids & bases and cursing my lack of editing skills! The (relatively) simple version is that pH is a measure of the concentration of hydronium (H3O+) ions relative to hydroxide (OH–) ions in water. In a neutral solution like pure water, they are at equilibrium and there is the same amount of each. The chart below shows the different ratios of hydronium to hydroxide ions at each pH. You will notice that in the red section there are more hydronium than hydroxide – this is acidic. In the blue section there are more hydroxide and less hydronium – this is alkaline (aka basic).
pH is mainly a useful way of describing a chemical environment, as it helps to explain how other chemicals will react in that environment. For example, when a low pH (acidic) solution reacts with many metals, hydrogen gas and a metal salt are created.
pH is one of the fundamental attributes that affects living things – including plants. In living cells a difference in pH across the cell membrane is harnessed to drive some of the most fundamental processes for life itself – photosynthesis and respiration.ref1,ref2 Living things are generally very good at managing the pH inside their cells and have feedback processes to adjust it up or down according to their needs and the environment (called homeostasis). Studies have shown that pH within plant cells is maintained at a small range of 7.1–7.5.ref
It’s when plant cells interface with the outside world, such as when taking in nutrients from the soil, that pH can make a difference to the efficiency (or not) of these reactions. Nutrients are taken up by plants as ions – ie. dissolved in water. This means that they need to be in solution for root hairs to take them up, and that solution can be acidic, alkaline or neutral.
Dissolved substances in the soil water (which change its pH) can also change the availability of nutrients – for example calcium ions will react with phosphorus ions to make calcium phosphate, so the phosphorus is unavailable for plants.ref But plants adjust their uptake according to these changes, so when they detect pH levels which reduce nutrient availability, in many cases they adjust their uptake to compensate, and these forces work in opposite directions.ref The overall effects of pH on the availability of nutrients to plants are a combination of the effects of pH on absorption by soils and the effects of pH on plant uptake.
Below is a chart showing the absorption of different nutrients by soil (in this case geothite, an iron rich soil). You can see that due to their different chemical makeup, each nutrient has a different absorption rate – the higher the absorption, the less available for plants.
Negatively charged metals (‘anions’) have a more consistent soil absorption profile – and most are absorbed by the soil eventually when the pH is 6 or above. But uptake by plants is significantly increased as pH rises.
So far it seems like acidic soils might provide more nutrients – but also more toxins (eg. cadmium, lead & aluminium). But the release of organic matter, including nitrogen, sulphur and the activity of microbes which perform this breakdown, is increased at higher pH, and the uptake of metals is increased.ref So it’s really a conundrum to work out the net effect of all these interactions! What do we actually know? Some findings include:ref
Phosphate fertiliser is least effective near pH 7; it is necessary to apply more of it to achieve the same yield as at lower pH. It is most effective near pH 5
Boron uptake is consistent between pH 4.7 and pH 6.3, but a 2.5-fold decrease occurs at pH 7.4
Molybdenum uptake is eight time higher at pH 6.6 compared to pH <4.5ref
Uptake of metal ions from solution by plants is increased by increasing pH – but their availability is decreased. This applies to toxins as well as nutrients. Magnesium and potassium are two important nutrients to which this applies.
Sulphate’s absorption by soil decreases markedly with increasing pH but plant uptake also decreases – the net effect has not been determined.
There is actually a fantastic diagram which shows the best soil pH range for each plant nutrient – you can see this all over the internet and it looks so useful! But unfortunately this diagram, which was created in the 1940s, is incorrect and has no real numbers behind it.ref In reality “nutrients interact and different plants respond differently to a change in pH” as described above so there is no one-size-fits-all diagram.ref
While I’m in mythbusting mode, there isn’t any such thing as ‘soil pH’ either! As noted in this excellent study from March 2023, pH can only be measured in a liquid. Unless you are over-watering, it’s likely your soil is not a liquid, therefore the soil itself does not have a pH. The pH that is being measured when ‘soil pH’ is measured is actually the pH when the soil is mixed with water – whilst this is indicative of the pH that might be present on individual soil particles, there is probably a range of pH instead across different particles. The pH of the water on a soil particle and the pH of the water on a root hair combine to create the true pH environment for a particular nutrient on a particular root. This is obviously not very easy to measure! See the end of this article for my bonsai media pH experiment.
The study mentioned above basically claims that most studies on pH and soils have failed to take into account the interplay between availability in the soil and plant uptake of a nutrient, which often work in opposite directions and so pH should not be taken to be the main factor in nutrient uptake except in specific circumstances. But looking at all of the above, it does seem like slightly acidic conditions should optimise all of the different reactions taking place – between 6 and 7 pH.
To bring it back to my bonsai, in my London garden with hard tap water of pH near 8, on the surface it would appear that this has the potential to cause a phosphorus deficiency in my plants, and perhaps affect their boron, molybdenum and metallic ion levels (we care about magnesium particularly which is used for photosynthesis – magnesium uptake increases at high pH but availability in the soil decreases).
But tap water is not the only thing affecting pH in the water in my bonsai soil. It’s also affected by the pH of my rainwater, which was 5.89 on the last measurementref, as well as the medium in my pots. I use composted bark, biochar and molar clay. Composted bark has organic components so is acidic, biochar is slightly alkaline and molar clay appears to be acidic – and this pH will become evident when particles of these components dissolve into the water. So the actual pH of the solution in my bonsai soil is anyone’s guess! All I can conclude from this is that a long summer without rain might cause my soil to increase in pH due to the removal of one acidic component – the rainwater.
The other thing to consider is that you can obviously adjust the availability of nutrients by adding them to your soil. So even if uptake is reduced by a particular pH, making more nutrients available could compensate for this. Hence the importance of regular fertilising for our bonsai, and using a range of different fertilisers which provide different nutrients.
Finally if you want to test the pH of your bonsai medium, a good approximation can be made by using a red cabbage and some distilled water (don’t use tap water, as this will affect the outcome if it’s not neutral to start with). Simply boil up a bit of red cabbage in (distilled) water, let it cool and while you are doing that put a representative piece of your bonsai medium into some water (also distilled). Allow them to soak for a while. Remove the cabbage from the cabbage water, strain the medium out of the bonsai medium water, and pour some of the cabbage water into the bonsai medium water. It should change colour according to the pH as follows (you can read more instructions here):
I performed this experiment on different bonsai mediums I had sitting around in my shed by soaking them in filtered water for 1 hour, then adding the cabbage indicator. The results were interesting! I was expecting the Kanuma to be acidic but it was actually neutral, as was my bonsai mix (which included some molar clay, bark, biochar, pumice and compost), and the pumice was surprisingly slightly alkaline. A rather small amount of biochar caused the indicator to go dark blue, which definitely tells me it needs to be used in moderation (although other mechanisms in biochar make nutrients available to plants, which you can read about in my biochar post).
What I conclude from all this is that my use of composted pine bark in my bonsai mix is probably a good thing as it will counteract the alkalinity from the tap water. This was a suggestion I learned from Harry Harrington’s website – although he recommends it for water retention, it would appear to balance a high pH medium or water as well. It also has the added benefit of being organic matter, which is a fertiliser in itself, creating more nutrient availability even if the calcium carbonate in my water locks some away. The need for applying fertiliser regularly is also apparent, as you just don’t know how nutrients are behaving in your particular bonsai soil and you need to give each tree every chance they have to access the nutrients they need. But overall other than causing annoying limescale marks on pots, my bonsai seem completely fine with hard water.
If you’re a fan of Gardening Australia as I *massively* am, you will have noticed they are always going on about mulch. Mulch (often in the form of bark or woodchips) gets added religiously to everything they plant whether in a pot or in the ground. This got me wondering whether mulch could be beneficial for my bonsai.
What is mulch? Well back to my expert source Gardening Australia in their article Mulch, mulch, mulch, it is a layer of materials such as compost, bark and woodchip products, and/or various grades of pebbles and gravels which are placed on the soil. The benefits they claim for mulch include water retention, weed control, protection from extreme hot or cold, reducing erosion, delivering organic matter and nutrients into the soil, and even – that it looks good!
Actually I want all of these things for my bonsai, so what does the science say about the effects of mulch?
The main benefit most studies seem to agree on is that mulch reduces weeds, and the thicker the mulch the more weed reduction.ref In one study on container-grown Thuja plicata it was as effective as chemical weed control.ref This finding is repeated across many other studies as well.
How about reducing hot root temperatures? Potted tomatoes with grass mulch showed a direct relationship between mulch depth, soil moisture and soil temperature (see the chart below.ref Moisture was increased and temperature decreased with additional depth of grass mulch. I don’t think it’s realistic to add 10cm of mulch to a bonsai pot though!
In a winter study, chopped newspaper as well as other mulches moderated cold temperatures.ref The Thuja plicata study by contrast found no soil temperature improvement by using mulch, and they blamed the colour of the pots (black) for this.ref So it looks like there might be a positive effect on root temperature but not if you have black pots – and only if you put a decent amount of mulch on the soil.
What about water retention? A study using plastic mulch (ugh) on Japanese privet plants found that the water that needed to be applied was 92% less in mulched potsref but the Thuja plicata study stated that no change in water retention resulted. The researchers proposed that transpiration was the main driver of water use (and since this happens at the leaf surface mulch will not impact it).ref An intriguing study in South Africa found that only a mulch of white pebbles was useful for water retention in the hot summer, but mulches of other organic types (bark & leaves) were also effective at reducing evaporation during the colder winter period. They were pretty brutal with their research subjects – potted Polygala myrtifolia – which only got a watering once at the beginning of the trial and then had to tough it out for 6 weeks without any more water being added! In the summer period of the trial only 7% of survived, and 50% of these had white pebble mulch. During the winter trial 92% of plants survived and in these circumstances mulch of any kind provided a 20% improvement in soil water content relative to no mulch.ref So it looks like mulch provides some improvement in water soil content as long as it’s not a drought scenario (and you don’t have black plastic pots).
One thing I have noticed is that a layer of Melcourt propagating bark (2-7mm) on my bonsai seems to ‘suck’ the water into the pot in when I am watering. Several studies have found that a layer of mulch on soil increases water infiltration rates.ref1, ref2 This may be because the pieces of mulch are “able to absorb the kinetic energy of rainfall…[or watering]…and maintain soil aggregates longer” and result in “an increase in the tortuosity of water pathways due to the higher roughness”. A study on Holm oaks found that rock fragments were a good mulch for shallower root systems and improved soil moisture.ref A rough-textured mulch might be useful if water is bouncing off the surface of your planting medium.
So should you use some form of mulch on your bonsai? If you want weed control, probably. If you have trees which are particularly prone to drying out or succumbing to the elements – for example they have very shallow or small pots, or are potted in medium without some form of water retention (such as coconut coir, vermiculite, bark or sphagnum moss) it might be worthwhile. It may also act like a form of insulation (as discussed in the post on frost) to protect roots from the cold. Finally if your medium doesn’t want to cooperate with the watering can or hose, and water bounces or flows off it, mulch might be a way to reduce overflow and improve infiltration.
What options are there for bonsai mulch? There are quite a few different types of mulch described in this article but not all of these would be practical for a bonsai pot, and many you wouldn’t use for aesthetic or ‘aromatic’ reasons. Only a mulch with a relatively small component size would be feasible – this could include a small-sized bark mulch, or even a layer of smaller medium such as akadama, pumice or molar clay. I’d love to be able to use seaweed but I don’t think it would smell good, and it’s not that easy to find in suburban London. Organic mulches will break down over time and add organic matter to your soil – which you may or may not want to do. So – maybe this is a practice you might want to consider.
You’ve probably heard the term ‘nitrogen-fixing’ – it means extracting nitrogen from the air. Which doesn’t seem like it should be too difficult, since nitrogen makes up 78% of airref, but in reality plants can’t use gaseous nitrogen.
In nature (ie. where nitrogen is not added artificially as fertiliser) plants mostly rely on microorganisms to help them get nitrogen – they access it in dissolved inorganic forms as ammonium (NH4+) and nitrate (NO3-). This is the nitrogen cycle, where organic nitrogen from dead organic matter is converted back to inorganic nitrogen as ammonia (NH3), then ammonium, then nitrate.ref Although this is performed by a range of different bacteria and fungi, this is NOT nitrogen-fixing, it’s ammonification followed by nitrification.
Nitrogen-fixing is the specific act of extracting nitrogen from the air, and it’s also performed by a range of different bacteria, known as diazotrophic bacteria. Certain plants create symbiotic relationships with these bacteria, with the most effective being root nodule symbiosis. These plants have evolved to provide a safe home for nitrogen-fixing bacteria in their roots, in small nodules where the bacteria live. The bacteria get food from the plant and protection from the outside world, and in return the plant gets nitrogen. Plants which can do this all belong to the ‘FaFaCuRo’ group – Fabales, Fagales, Curcubitales, and Rosales – they are all flowering plants (angiosperms).ref You can download a database of all 825 known species with root symbiotic nitrogen fixation here – they include green manure such as clover and legumes, as well as some trees – Acacia (wattle), Casuarina (sheoak), Albizia (Persian silk tree), Robinia (locust), Wisteria, Alnus (alder), Elaeagnus (oleaster) and Hippophae (sea buckthorn).
The initial question behind this article was me wondering whether planting clover or similar nitrogen-fixing plants in my bonsai pots would achieve anything – like somehow supplying my tree with a free source of nitrogen. After looking into it further I concluded that the answer is no! Nitrogen-fixing plants have a great system – for themselves. The reason why they are used as green manure, or as rotational crops, is because they don’t require (or require less) supplemental nitrogen, so the land where they are planted gets a break from fertilizer. When they are harvested they can be ploughed back into the ground for bacteria to break down via ammonification/nitrification, so the next crop can benefit from a nitrogen source which hasn’t come from fertiliser. Basically it’s a way of making natural fertiliser – effectively compost – which hasn’t had added fertiliser as an input.
You could benefit from nitrogen-fixing plants such as clover for your bonsai practice – if you composted it and used it as organic matter in your soil mix. In fact it has been found that nitrogen-fixing trees in a tropical forest inhibit their neighbours (possibly due to their stronger growth rates), so you definitely don’t want your trees to share a pot with these species while they are alive.ref
There is also what’s known as ‘associative nitrogen fixation’ – this is when a nitrogen-fixing bacteria ‘associates’ with a species of plant without actually taking up home in root nodules. They are found on the roots, in the rhizosphere, and sometimes within plant tissues as endophytes.ref It has been suggested that up to 24% of nitrogen supply to cereal crops such as maize, rice and wheat is actually supplied in this way and that ‘mucilage’ (sugar exudates from roots) may be responsible for attracting the responsible bacteria.ref Although interestingly it may not actually be that the bacteria provide nitrogen directly, but instead they influence the plant to be able to access more nitrogen in the soil, for example by increasing root hair surface area.ref This is the mechanism by which biochar improves nutrient acquisition as well – by increasing the plant’s Nitrogen Use Efficiency or ‘NUE’.ref1,ref2
Which unfortunately brings us back to needing a source of nitrogen in the soil in the first place. What I have concluded is that unless a bonsai tree is a nitrogen-fixing species itself, the only way for it to obtain nitrogen is from the soil via the nitrification of dead organic matter, or by adding chemical fertiliser. And from a sustainability point of view, using at least some dead nitrogen-fixing organic matter (such as legume plants) for composting may be best as this is net-positive for nitrogen, bringing previously inaccessible air-borne nitrogen into the soil (so – go forth and compost your legumes!)
The main impact you can have as a bonsai tree custodian (aside from providing a nitrogen source) is to improve your tree’s nitrogen use efficiency so it can gain the most from the nitrogen which is present. There are a few ways to do this. Adding beneficial bacteria to the soil provides the associative nitrogen fixing effects explained above, and keeping the pot at the requisite temperature, pH, aeration and soil water level that is attractive to these microbes is also a factor – although it’s hard to know exactly what these conditions are! Avoiding extremes is probably the best approach. Adding biochar to the soil is known to improve nitrogen use efficiency.ref Encouraging a high root surface area through root pruning and encouraging root ramification is another contributor. Finally, do not overfertilise, as this has the opposite effect on root ramification since nutrients are easy to find and roots do not need to increase their surface area.ref
At a recent club meeting (shout out to Twickenham bonsai club) the subject was azaleas. During a critique session it was noted that on Satsuki azaleas, different coloured flowers appear on different branches so you need to be careful when you prune not to remove the colours you want to keep. This got me wondering about Satsuki azaleas, why they have the colours they do, and what this might mean for their bonsai custodians.
Starting from the start, flower colour is the “result of pigment molecules accumulating in cells”ref and depends on which pigment molecules are present, where they are distributed, and the shape of the host cell.ref
There are several different types of molecules responsible for colours in flowers – the main ones are carotenoids, flavonoids and alkaloids. There are 700 known naturally-occurring carotenoids,ref including substances in the carotene and lutein families like beta-carotene which is in carrots and lycopene which is in tomatoes – these produce yellows and some oranges. Flavonoids produce the widest spectrum of coloursref, and these include anthocyanins which make pink to blue-violet colours, chalcone/aurone which make deep yellows and flavones/flavonols/flavanones which make white and light yellows. Betalains don’t get as much press as the previous two pigments, but these are responsible for reds and deep purples (such as in beetroot) – and interestingly they never appear in the same place as anthocyanins, so it is either one or the other in a given flower.ref
In the case of azaleas, anthocyanins and flavonols are the major pigments.refOne study analysed azalea colours by their pigments, and showed that the different colours fit into three pigment groups – red, purple and white. They found that the red group contained two to four major anthocyanins, and the purple group had two to six – therefore more colour variation is possible in the purple group. They found also that white group flowers did not contain any anthocyanin, but did contain the precursors to this pigment, so their lack of colour is probably a genetic defect in the pigment biosynthesis pathway which causes pigment creation to fail.
D. Mizuta et al. / Scientia Horticulturae 122 (2009) 594–602
Pigment molecules are what are known as ‘secondary metabolites’ in plants – substances which they synthesise to help them in some way or other.ref Aside from those mentioned above, chlorophyll is another pigment that plants make, this appears green and is responsible for photosynthesis. Without going into the detail of the chemical structure of each pigment, the key point for this article is that plant cells need to synthesise a pigment or usually a combination of pigments in order to display a particular colour. The synthesis of pigments is a multi-stage process which is mediated by enzymes, and the production of enzymes is under genetic control (see a cute animation of this here). So genes – contained in the DNA within chromosomes in every cell – determine what pigments are produced where when cells develop. To learn more about the biosynthesis pathway for azalea pigments you can read this paper (a reasonable understanding of genetics is needed to make sense of it). This seems like a good moment to state that I am not a geneticist! So the below is my citizen-scientist interpretation of what I have read – if there are errors I would be happy to correct them.
The most obvious phenomenon illustrated in these examples is that each plant develops flowers of different colours and patterns. Some flowers are one colour (red), some another (white), some have different coloured petals, some have coloured stripes and others have speckled colours. Not shown here are the different flower shapes, which can also be different on the same plant. Each one of these ‘phenotypes’ (observable traits) has its own explanation.
The ability of a single plant to produce different phenotypes for flowers is known as ‘sporting’ and the branches which have this variation from the ‘base’ colouring are known as ‘sports’. Sports result from “sudden variations in gene expression of somatic cells…[which]…results in plants having a different phenotype” – in plain English this means that one or more cells which produce a shoot or flower bud all of a sudden start to use different instructions for creating pigments.ref According to an excellent South African study by S de Schepper et al, which I have used extensively in writing this post, “all the information required to sport (the ‘sporting capacity signal’?) is present in leaves”, which suggests the capacity for sporting is across the tree – it’s a characteristic possessed by the plant as a whole, which is triggered in a specific cell as a ‘sporting event’.
How a sporting event happens is that the meristem (the growing tip of the plant) spontaneously develops a mutation in a cell, and every cell that divides from this cell carries the same mutation. The South African researchers investigated this for azaleasref and found that the majority of these changes in azaleas are ‘epigenetic’ events such as those caused by ‘transposons’ or so-called ‘jumping genes’ref. These types of sports can only be passed on to new plants by clonal propagation, since epigenetic events are by definition not captured in seed or pollen.
But what is clearly passed on genetically is the ability to produce sports in the first place, this is obvious by the fact that so many Satsuki azalea varieties which are created by sexual reproduction have sporting behaviour. Probably the best hypothesis I have come across to explain this is that polyploid parent plants are partly responsible. Mark Nijland notes in an article in 2022 that ‘Suisen’ is a dominant parent plant in Satsuki breeding.ref It turns out that ‘Suisen’ is not only a tetraploid (ie. it has four sets of chromosomes instead of the normal two), it’s also a mixaploid, which means it has different genotypes in different parts of the meristem – some areas are diploid (with two sets of chromosomes) and some are tetraploid (four sets of chromosomes).ref Other tetraploid/mixaploids include ‘Shinsen’, ‘Miharu’ and ‘Koyo’ and there are also a number of known Satsuki azalea triploids (having three sets of chromosomes).ref What polyploidy/mixaploidy brings to the plant is a larger number of options (known in genetics as ‘alleles’) for development – so that when a flower is created it can have multiple different colour or pattern options.
Another factor is that polyploid organisms have to deal with what is called ‘genomic shock’. When more than two sets of chromosomes are combined in a single cell there is more potential for abnormal development, which can negatively affect an organism’s chances of survival. Plants are known to use transposons to adjust their genes to avoid this genomic shock.ref (This is usually the moment in a research paper or article where the author calls out the remarkable Barbara McClintock for her discovery of transposons, she also proposed the idea of genetic shock. She presented her work at a symposium in 1951 to great derision and wasn’t believed for over a decade – her Nobel Prize wasn’t awarded until 1983.)
Anyway, transposons jump into a gene and change its function – switch it on or off or cause it to do something different. Due to managing genetic shock a polyploid plant is more likely to be making use of transposons than a genetically stable diploid (the ‘normal’ two sets of chromosomes) plant.
The theory goes that in Satsuki azaleas, the presence of polyploidy and mixaploidy result in plants which are more likely to spontaneously adjust their genetic expression via transposons, and when this happens, a wider range of colour and form options are available due to the larger set of genetic material from which to choose.
A prevalence of polyploid/mixaploid parents would undoubtedly be down to selection by Japanese breeders over the centuries. Azaleas have been intensively hybridised and selected since the 1600s, and in 2020 there were 1400 varieties listed in the Japanese Satsuki dictionary.ref The polyploid parents such as ‘Suisen’ must have been observed to produce sporting child plants over the long term, and were retained for breeding (this article notes that Suisen is the most dominant parent over three decades of cultivar production). It has been found by genome analysis that not only did the ancestor of azaleas have a whole genome duplication event 75 million years ago, but also that the genome of Rhododendron simsii, a parallel branch of rhododendron to Satsuki azaleas (which are mainly Rhododendron indicum) is 47.48% comprised of repeat elements, the majority of which are transposable elements (transposons).ref So it’s not surprising that somewhere along the way azalea breeders came across varieties which dialled up the ‘sporting’ aspect of their genetic makeup.
Let’s take a look at some of the different types of sports which are observed on Satsuki azaleas and why they form. First we need to understand one key attribute of sporting events which took me several days to get my head around – once you do it is a bit of an ‘a-ha!’ moment. This key insight is that sporting events can happen anywhere on the meristem – for example half-way up a shoot as the meristem is happily dividing – they don’t necessarily neatly happen just as a flower or shoot bud is emerging. Wherever a cell is dividing and generating a new cell, a sporting event can take place. If a sporting event related to flower pigment happens halfway up a shoot, you won’t notice it until a flower emerges but that sport will be there on the stem and on any leaves which bud from that stem. So to understand how flowers on a stem might be affected by sporting events you need to imagine the time-sequence of your azalea developing and how a sporting event would propagate along a stem as the plant grows.
Different but uniform colours on each flower occur when a sporting event changes the gene expression for pigments across the entire flower – so all cells are affected. For example, a pigment isn’t expressed any more, or a pigment which was suppressed is now expressed. It is said that darker colours in azaleas dominate geneticallyref. So in the image above, the ‘cancel pigment’ genetic defect was already present in the tree – to produce white flowers. As the tree grew and new stems developed, some transposons jumped into the defect in a cell which was a precursor to an entire stem or branch, and deactivated it. From then on the flowers would be red due to the red pigment being produced. Some branches haven’t had the transposons jump yet, so they stay white.
Stripes are caused by ‘genetic mosaics’ or chimeras where different cell layers or areas of cells within the meristem have different genotypes. So part of the stem where this flower bud developed has the sport and part does not – and both of those sets of cells contributed to this flower. In the example above you can see a flower with a clearly delineated section of dark pink on the left. This is an indication that the sporting event (in this case reactivating the pigment production) happened at a point which you can trace back along the two edges of pink. These flowers may be relatively solitary on the tree, since it’s their unique position and timing of development which gives them these patterns. You can imagine a small stripe of pink along the stem where this flower bud came from, back to the location of the sporting event, and one might continue along the stem above this flower as well.
Flecks are another version of chimeras but the mutation is not persistent in the same way as stripes – you could call them ‘micro-sports’. One explanation for this behaviour could be that the transposon which reactivated the red pigmentation ‘flickered’ on and off as the flower developed. Unlike stripe chimeras, flecks can be passed down to child plants, indicating that ‘flickering’ behaviour is a genetic trait and not an ephemeral one like most other sports.ref
Petal borders or margins of a different colour on the same flower (also known as picotees) can arise in different ways. In flowers with large margins (7mm or more) this is due to different cell lines being mixaploid, resulting in the petal edges containing 4 sets of chromosomes and the rest of the flower containing 2 sets of chromosomes. In one study this made petal edges white and the centre red as in the picture above.ref Other picotees are caused by ‘positional’ differences in gene expression where all cells have the genes necessary to make pigments, but only those in a specific position are ‘instructed’ to do so.ref In both cases these are due to somatic/spontaneous changes and cannot be passed down through sexual reproduction.ref
Combinations of effects are seen on many Satsuki azalea varieties – the one above has stripes, picotees, flecks and three different colours – intense pink, mid-pink and white. Having all these effects together comes down to the mastery of the breeder, in combining the right parents.
Effectively Satsuki azaleas are like giant 4-dimensional genetic puzzles, with gene switching happening all over the plant at different moments in time to create a completely unique plant! No wonder people get obsessed with them. So finally, if you’ve reached this far through my total geekfest immersion into Satsuki azaleas and their genetics, what does it mean for bonsai?
Well, hopefully the above will help you understand that ‘sporting events’ or spontaneous mutations are what cause many of the different flower colours and patterns seen on Satsukis. And that the type and duration of the sporting event will determine what colour/pattern change happens upstream. You might also be able to work out roughly where these events have taken place on the plant which will allow you to trace where they will go in the future and to either encourage that growth or prune it away, depending on your goal. This information might also help you choose the right variety if you’re buying a Satsuki azalea – especially if you want lots of variation, choose a polyploid/mixaploid or one of its children.
A final question I had for this article was if there was any way to increase epigenetic events to generate more variation on the plant and increase sporting. The answer is yes. “Stresses such as wounding, pruning, viral infection, and tissue culture are all known to induce the movement and/or activity of Transposable Elements, as well as DNA damage”ref One relevant stress in our case is pruning. Pruning back hard (as is advocated by azalea specialists such as Peter Warren of Surayama Bonsai) should generate variation since it creates lots of new growth (ie. cell division and opportunities for mutation) as well as a certain level of stress. DNA damage is another one, which can be caused by exposure to ultraviolet light.ref So in theory keeping your azaleas in full sunshine could also encourage sports. However note that as mentioned above pruning back hard might have the effect of removing a sport! If you have a really excellent sport, make sure to note where it is to avoid removing it.
I want to mention one other resource I used for this article – there is a BonsaiNut member – Harunobu – who although not recently active appears to have excellent understanding of the genetics of Satsuki. Some of their advice I found interesting although I could not verify it through other means.
Aside from confirming the theories above about transposons causing sports, Harunobu says that in azaleas “colour is dominant over whiteness, intense colour is dominant over paleness, purple is dominant over red, and solid coloured flowers are dominant over bicolour/variegated flowers”.ref This means that over time, as the meristem expands and divides, the dominant colours or regions may start to – well – dominate. That implies that assuming you like the less dominant characteristics, you should cultivate them as much as possible and not remove or dilute them. Similarly the more dominant characteristics should be pruned back to avoid them taking over all the subsequent growth. Of course – a new sport can arise spontaneously anyway to give you something new to work with.
Another comment from Harunobu was “a tip that might be marginally useful for when you are trying to ID an azalea by flower colour when it is not actually in flower. If the old leaves go yellow: it is white flowering. If the old leaves go orange; it is pink. If they go red; it is red or purple. And if they have sectors on the leaves, they have sectors on the flowers.”ref I assume this is because the pigmentation in the flowers also carries through to the leaves and when the chlorophyll dies away the pigments become visible. Theoretically you might be able to use this to visualise sports on a plant without it being in flower. However when I tried this on two of my own Satsuki, I could not see sectors (stripes) on the leaves, and all the old leaves were yellow although the trees are mainly pink. So – see if this works for you.
A weird idea I came across researching this article was the idea that somehow dominant characteristics could ‘take over’ a plant and actually influence branches which previously had been different colours or patterns. I don’t see how characteristics can literally go backwards and influence cells which are behind them in the developmental sequence. And surely new sports would appear? I can’t explain this idea and don’t think it’s true (however, I am open to being wrong). The only way I can see for this to happen is if you don’t prune at all and let everything grow out, then maybe it might be the case that the dominant characteristics take over all of the new growth.
I’ve talked about transpiration in quite a few different posts on this site, but a recent thread on http://bonsainut.com caused me to think maybe I should have a post dedicated to it, so here goes…
Transpiration is the evaporation of water from the leaves of a tree. It’s actually a critical process for trees, because excess transpiration is one of the few ways in which a tree can die; so-called ‘hydraulic failure’ has been identified as the most prominent cause of tree death.ref Hydraulic failure – the failure to access enough water to replace water lost mainly through evaporation – causes cell death, xylem failure and a fatal reduction in photosynthesis. So it’s really important for bonsai practitioners to understand this process.
The main driver of transpiration is not – as you might think – to cool the leaves (although this is one reason for it). In fact transpiration is a by-product, or ‘cost’, of photosynthesis, and it happens because of the way that leaves obtain carbon dioxide. You may already know that plants have small pores called ‘stomata’ which open up to let air – and CO2 – inside the leaf. But you might not have known that gaseous CO2 in air needs to be dissolved in water before it can be accessed by chloroplasts and used for photosynthesis (this is explained in Vogel (Chapter 5 – ‘Leaking Water’). This means that water needs to be available on the surfaces inside the leaf – which means that when stomata open up, this water is subject to evaporation.
Vogel says that “only if the relative humidity is 100 percent will water not be lost…[and]…if the leaf’s temperature is above that of the surrounding air, then water can be lost even at that humidity.” He also says that for every gram of CO2 used by a leaf for photosynthesis, it’s estimated that 125 grams of water is lost.
Smith et al (Chapter 4.10 Movement of Water & Minerals) explain that this evaporation causes a constant flow of water known as the ‘transpiration stream’. As water evaporates from the leaf cells, pressure in those cells is reduced, and this negative pressure causes water from the xylem to move into the space, due to strong mutual attraction between water molecules. That in turn pulls more water behind it and so on. This hydraulic mechanism is responsible for pulling water all the way up the tree from the roots. Actually this process is fundamental to the health of the tree, maintaining cell turgor (stiffness), transporting nutrients, metabolites & growth substances synthesised in the roots throughout the tree, and providing a source of water for the phloem stream which flows in the opposite direction providing energy to the tree. When there is enough water available, all of this works perfectly – when there isn’t, problems arise.
The extent of evaporation from the leaves of a tree is determined by several different factors, which can be divided into environmental, tree-specific physical factors and tree-specific response factors.
The main environmental factor which drives transpiration is the ‘vapour pressure deficit’ (“VPD”) – this is the “difference between the amount of moisture in the air and how much moisture the air could potentially hold when it’s saturated.”ref VPD is a function of both heat and humidity, and provides a measure of how powerful the evaporative force of the air is with any combination of these.ref
Occasionally while writing articles for this blog, I end up in the world of cannabis cultivation. Maybe because they are very motivated to keep their crops vigorous, cannabis growers and their equipment suppliers sometimes have the best data and charts out there! This is just such an occasion, see below for an excellent chart from ‘Ceres Greenhouse Solutions’ showing the VPD for a given temperature and humidity (you can download a copy here). The VPD is low in the blue section and high in the red.
What you will notice is that the relationship between humidity and temperature isn’t exactly linear. Also, VPD increases with higher temperature and lower humidity. Since a higher vapour pressure deficit means there is more ‘pull’ on the water in leaves, increasing temperature and decreasing humidity both increase transpiration – and they reinforce each other, so dry and hot is a high transpiration combination.
Another environmental factor is wind. One study found that wind actually improves water use efficiency, because whilst it does increase transpiration, it also increases CO2 uptake, and the net effect is greater water use efficiency and not less.ref But for the purpose of this article, wind does increase transpiration.
Coming onto tree-specific physical factors – these are all the attributes that relate to the size, shape, position and structure of the tree. In general the more foliage a tree has, the more it will transpire – so a large broadleaf tree will transpire significant amounts on a hot day – in one study they found a large canopy tree in the tropics (Eperua purpurea) transpired up to 1180 litres per day!ref By comparison in the same study, smaller (presumably more shaded) trees transpired a lot less. Thomas (Chapter 2: Leaves the food producers) gives the following figures: “<100L/day in conifers, 20-400 L/day in eucalypts and temperate trees such as oaks, reaching perhaps 500 L/day in a well-watered palm and as high as 1200 litres per day in specimens of Eperua purpurea growing out of the top of the Amazonian rainforest canopy.”
The chart below shows the daily transpiration rate during the growing season for a sessile oak tree in Turkey which measured 18.5m x 34.5m – this maxed out at 160 kg/day (effectively 160L).
As well as the volume of foliage, trees have different stomatal size and density (number of stomata in a given area) which are determined by genetics as well as environmental factors (such as intensity of light and VPD to which they are exposed when developing).ref1ref2 Low stomatal area (ie. density x size) will result in lower transpiration when compared to a tree with higher stomatal area. These researchers measured stomatal area for 737 plant species across 9 forests and at the lower end of the spectrum conifers such as Cunninghamia lanceolata (0.2%) and Picea koraiensis (0.4%) had 100 times less stomatal area than angiosperms such as Viburnum betulifolium (23.77%) or Quercus serrata (21.74%). You can download all their data here. Basically the more stomatal area which is open to the air, the more transpiration there will be.
Many trees have wax plugs in their stomata which reduce their efficiency, and transpiration at the same time. To copy a piece from my article on needle leaves, wax deposits in Sitka spruce stomata reduce transpiration by two thirds but photosynthesis by only one third.ref One study found that 81% of the species they looked at contained such plugs and that wax plugs are particularly numerous in conifers.ref
Another factor is the level of transpiration via bark. This isn’t due to stomatal opening but simply due to partial permeability of bark to air – also genetically determined and due to the presence of ‘lenticels’ – small channels which allow passage of water and air for the metabolism of living cells in the bark. One study on Pinus halupensis found that “Bark transpiration was estimated to account for 64–78% of total water loss in drought-stressed trees, but only for 6–11% of the irrigated trees.”ref This is because bark transpiration is passive and unmanaged, unlike leaf transpiration which can be somewhat controlled by the tree (see below).
Also relevant for individual trees is their position relative to other trees and the sun. Shade will reduce the temperature at the leaf surface and reduce transpiration, a mass of trees together along with undergrowth may increase humidity, also reducing transpiration. A tree standing alone or above others will be exposed to higher temperatures and lower humidity, thus increasing transpiration. Different areas on a single tree will be exposed to different combinations of these factors as well, so rates of transpiration will differ even from leaf to leaf on a given tree.
The final category of attributes which determine transpiration relates to the trees’ ‘behaviour’. That is, how they react to different environmental conditions. As we all know trees may be sessile but they are also incredibly dynamic and can adjust a wide range of parameters of their own biology. The main issue they need to address in this case is losing too much water, which could lead to death. As a result, they change their physiology to manage evaporation as well as water intake at the start of the transpiration stream.
To manage evaporation, trees adjust their stomata based on water availability, changing their ‘stomatal conductance’ to reduce transpiration if not enough water is available.
They do this in a couple of different ways – ‘passively’ and ‘actively’.ref The passive mechanism is where lower water pressure within leaves causes guard cells around the stomata to lose their stiffness, which has the effect of reducing the stomatal aperture. The active mechanism relies on the tree producing abscisic acid (ABA) – this “triggers efflux of anions and potassium via guard cell plasma membrane ion channels, resulting in decrease of turgor pressure in guard cells and stomatal closure”.ref
A study on Metasequoia glyptostroboides found that in most conditions of water availability the passive mechanism was in play, and it wasn’t until prolonged or severe water stress was experienced that the active ABA-mediated mechanism came into play.ref The article explains that different gymnosperm species use different combinations of these passive and active processes to manage a lack of water availability by reducing transpiration. Angiosperms by contrast use a more sophisticated and more recently evolved version of the active process, mediated by ABA.ref
Thomas says that stomata usually close when it is “too cold or dark for photosynthesis” or when the leaves are in danger of losing too much water and wilting”. The consequence of stomatal closing is an associated reduction in photosynthesis – so when a tree is drought stressed, it won’t be generating energy at the same rate as when it was healthy. A study measuring photosynthesis versus stomatal conductance for Pinus radiata (see in the chart below) found there was a roughly linear relationship, as the stomatal conductance increased, so did photosynthesis.
There are several other ways that trees manage their transpiration – by adjusting their root conductance (ability to draw in water), changing their leaf expansion so that there are fewer/more leaves which are smaller/larger in area, pointing exposed leaves downwards during hot periods of the day, changing the root/shoot ratio to match water source to water use and by operating a daily cycle of metabolism which optimises transpiration (eg. increasing their root hydraulic conductance at night when there is lower evaporation, and ‘filling up’ to deal with higher transpiration during the day).ref So they are very much active participants in responding to and controlled their transpiration rate.
But what does it all mean for bonsai? The first thing is, if your tree has plenty of water availability, transpiration should not become a problem, but you need to remember that up to 95% of water use is evaporationref so trees need a lot more water than you might expect. The best way to avoid issues associated with excess transpiration is to supply your trees with all the water they need. This is achieved by regular and sufficient watering, and by using a medium which has some water retention to avoid drought stress – but is also well-draining. A well-draining medium allows you to water more often without the risk of waterlogging roots or creating conditions for pathogens to take hold.
Also – a tree’s ability to handle water loss varies widely depending on the species – Thomas gives the examples of eucalypts and alder as species which cannot control transpiration effectively, and some oaks as species which can. So each tree in your collection will be different.
But let’s consider all the factors explained above that increase transpiration: high vapour pressure deficit (high temperature and/or low humidity), wind, lots of foliage, high stomatal area, clean (unwaxed) stomata, passive bark & leaf evaporation, a sunny/solitary/high position, and a lack of water availability to the roots which activate stomatal closure.
Some of these are adjustable for bonsai. If it’s going to be a hot, dry, windy day then your trees are going to transpire a lot more than normal and if their roots can’t keep up, you need to improve their environment; newly collected and recently root-pruned trees or trees in particularly small or shallow pots will be most affected. You can help them by providing shade (reducing the temperature), increasing humidity, and moving them out of the wind – and obviously by watering. For a temporary period, on a very hot day, it might even make sense to sit pots in water (do not do this for an extended period).
Transpiration can also be a problem in the winter as trees do continue to transpire, albeit at lower levels, even if they are deciduous. As such, they do need water to be available which means you need to keep an eye on moisture levels in pots. If they get dry, water them. If the medium is frozen, this will lock up water and can have a dehydrating effect so in this case you need to also water, ideally when it’s above freezing. Mulch is suggested to avoid hydraulic failure for trees in the groundref, a similar approach can be used for bonsai in pots, to reduce freezing and make more water available to roots. Even at night it is not the case that transpiration completely stops – typically it is 5% – 15% of daytime rates.ref
Balancing the amount of foliage with the roots when repotting or pruning is another important way to help your trees manage their transpiration rates, so that there is enough root mass to meet transpiration demands. Root pruning in the heat of summer should be avoided unless a comparable foliage reduction takes place. If you’ve gone a bit far with the root pruning, use the approaches above – provide some shade, increase the humidity and maintain a watering regime. This is where the bagging method for collected trees comes from – it reduces transpiration by increasing humidity and can be used for trees struggling to recover from a severe root prune.
Anti-transpirant is a product that some bonsai aficionados use. This does what it says on the tin – it is a “film-forming complex of polyethylenes and polyterpenes that when applied to foliage will reduce the moisture vapor transmission rate”ref The active substance is derived from conifer resins. In reducing transpiration these products also reduces photosynthesis, which is a consideration. I’m personally not a fan of disrupting a plant’s natural processes in this way, and successful use of the product depends on the individual tree and product selected (read more here).
Hopefully you can see from all of the above that transpiration is an extremely important concept to understand as a bonsai geek, but one which can be managed, as long as you are aware of the factors at play. Here’s to helping our trees avoid hydraulic failure!
Living in London means that even though spring has blossomed forth, there is still a chance of frost all the way through to the end of May. And many of us will have learned the hard way that frost and/or sub-zero temperatures can seriously damage our trees.
One of the key risks of frost and extreme cold is the problem of ice. Ice formation pulls water from plant cells, causing dehydration, which can be just as lethal as dehydration from underwatering.ref Ice masses which form and thaw can deform and damage cell membranes, as well as buds and other plant organs, and ice in trees’ xylem vessels can cause embolisms (air bubbles) to form, cutting off the water flow above the bubble.
Another problem is that low temperatures make photosynthesis dangerous due to an excess of energy which ends up as damaging reactive oxygen species (“ROS”). This is why some trees convert green chloroplasts to bronze chromoplasts in cold weather – read more here, and others shed their leaves altogether to eliminate the problem.
Plants which are ‘hardy’ or ‘frost-hardy’ have mechanisms to resist the effect of ice – either by ‘avoidance’ such as supercooling (lowering a liquid to below freezing point without freezing), or ‘tolerance’ – using biochemical changes and physical adaptations allowing the tolerance of ice in their tissues.ref Some pines have evolved specific mesophyll cells which allow the ice into the intracellular spaces without deforming the key structures within the needles – a physical adaptation. Oaks and ring porous trees regrow their conducting xylem every year, since embolisms during winter make last seasons’ xylem ineffective.
Crucially, since many cold-hardiness mechanisms rely on biochemical changes within the plant, they require time, so that the relevant proteins, enzymes and other metabolites can be synthesised in sufficient quantities to have the desired effect in plant cells. This process is called ‘cold acclimation’. Smith et al (2010) give the example of rye. 50% of non-acclimated rye plants will die at -6oC, but after spending 2 days at 4oC, they can go down to -21oC before 50% of plants will die. The cold acclimation process is what’s known as an ‘epigenetic’ process – where environmental triggers such as shortening days and reducing temperatures turn on genes – in this case known as COR (cold-regulated genes).ref These then produce the proteins which are used to create the cold-hardy biochemical changes.
There is an equal and opposite process known as cold deacclimation. Smith et al (2010) say that above a temperature of 10oC cold hardiness is rapidly lost, which is why a spring frost at the end of May can be so damaging.
To complicate matters, shoot hardiness is not the same thing as root hardiness.ref Studies have shown that roots are less hardy than shoots, even when exposed to identical temperature acclimation treatments.ref For example, the stems of Pyracantha are hardy to -25.6oC, whereas mature roots are hardy to -18.8oC and young roots are hardy to -6.1oC.ref This is an important insight for bonsai because the scale of our trees and the fact they are suspended in pots means their young roots are particularly vulnerable. A symptom of winter damage to young roots is when a tree flushes later than normal, and has retarded growth during the season.ref It may not kill the tree, but it will certainly give it a handicap for the next season.
So what does it all mean for bonsai?
For me the main risk boils down to root damage. Those lovely fine roots we work hard to encourage during the growing season are more vulnerable to frost than any other part of the tree, and unfortunately due to the size, shape and positioning of most bonsai pots, they are very exposed to cold temperatures. In one study it was found that container-grown trees were subjected to temperatures down to -15oC when the night-time temperature reached -30oC, whereas the soil only went down to -6oC.ref And since surfaces cool faster than air when it’s cold, resting up against a cold pot surface is the last place a young root wants to be! When the temperature gets really cold, mature roots can be damaged as well, which could be fatal to the tree.
Sure – leaves can be damaged by frost as well – mainly when deciduous trees have leafed out expecting above-zero temperatures and then a frost comes along. Most will deal with this and should regrow their leaves – there will be an energy penalty which will reduce the overall energy they have to devote to the growing season, and they won’t look great in the meantime, but it shouldn’t be fatal if the stems and roots are still healthy. But certainly to keep your trees looking and growing their best, you want to avoid deciduous trees leaves being exposed to frost if at all possible (see below for some strategies).
To be able to manage and prevent frost damage, we need to know when a frost will happen. This starts with monitoring the temperature during winter. Importantly, when you hear or see a temperature forecast for a location, it is a forecast for the air temperature. The temperature of the ground is often several degrees lower. The UK Met Office says “As a general rule of thumb, if the air temperature is forecast to fall between 0 °C and 4 °C on a night with little or no cloud and light winds, then you need to bear in mind there may be a frost outside in the morning. The closer it is to zero, the greater the chance of seeing frost. If the air temperature is forecast to be below zero, then the risk of seeing frost is much higher.”ref This goes for surfaces like pots as well. Wind and cloud cover reduce the chance of frost at a given temperature.
The actual temperature which will damage or kill roots is species-specific, so there’s no hard and fast rule but knowing where a tree comes from should give you an indication. In general the species common in the boreal forests such as many Pinaceae and Betula pendula will cope best with freezing temperatures. Some ornamental tree ‘killing temperatures’ are provided in this list.
So what should you do to help your trees defend themselves against frost damage?
The first thing to do is to let them be exposed to the cold over time, before it freezes. Give them time to activate their COR genes and establish cold acclimation. This includes not putting them in a polytunnel or shed or wrapping them up until they’ve had some time in colder temperatures (but not freezing).
It’s also useful to have actual temperature data from the location where your pots are going to be over winter. This can be achieved using a digital thermometer/hygrometer which records the temperature and humidity at regular intervals – there are many wireless/bluetooth-enabled options now which are very reasonably priced (I just bought 3 for £35, one for my garden bonsai bench, one for my allotment bed and one for my allotment greenhouse). This will enable you to monitor what the temperature does relative to the forecast so you can better predict the forecast in your actual location – and you can see which locations might provide better winter protection. If you see the temperature approaching zero you can act to protect your trees.
Then, adopting a form of overwintering system could be beneficial. Somewhat counter-intuitively, this involves watering everything well, without overwatering.ref The reason for this is to avoid dehydration. Then providing a physical barrier to the cold, effectively a ‘tree duvet’. This can move the frost surface away from the pot edges. You could use an insulating blanket of some kind, but do some research – one study found that ‘microfoam thermoblanket’ made a difference, but clear, black or white poly did not.ref The covering would need to have insulating properties, and not get saturated with water such that it would then freeze anyway. You could also move your trees into a polytunnel, garden shed or greenhouse, with the walls and air inside acting as a form of insulation. If you use a digital thermometer you can monitor these to ensure they aren’t getting too cold – note that polytunnels have been found to get cold enough to damage roots.ref
The absolute best is to provide some form of heat so that the temperature can’t go below a certain point – in practice this could be moving your pots closer to your house, using a (lightly) heated propagation bed, or putting them in a (lightly) heated greenhouse or outhouse. You don’t want them to think it’s spring so there shouldn’t be too much heat, just enough to keep the temperature above freezing. Be aware that in a warmer environment such as a shed your trees will deacclimate earlier as well, so make sure they don’t go back outside into a frost as they may have lost their cold acclimation.
Spring has sprung (finally) in the UK and now is the time that many trees flower, so they can pollinate the next generation with enough time for seeds to develop before winter. Plant reproduction is quite a complicated multi-stage process, but for the purposes of this post it’s enough to know that pollen is the vehicle for the male gamete (the plant equivalent of sperm). It is produced by anthers in angiosperms (flowering plants) and by male cones in gymnosperms (including conifers).
It turns out that pollen production can happen in different places. In some trees, both male and female parts are within the same flower (known as ‘perfect’ or ‘complete’ flowers). In others, they are separate flowers or cones on different parts of the same branch, shoot or tree – known as monoecious trees. In yet others they are separate flowers or cones on single-sex trees – known as dioecious.ref
Whilst female flowers/cones and the fruit associated with them are usually obvious, the pollen-bearing male flowers/cones are often hard to spot and sometimes people don’t even realise they are there (it was only about 2 years ago that I realised that oak trees even had flowers!). After releasing their pollen, male reproductive parts usually drop off and disappear from view, while the female flower/fruit/cone persists as the seeds develop. So it’s definitely worth appreciating male tree flowers when you see them, as they don’t hang around for long.
In angiosperms, many male tree flowers take the form of catkins. A catkin is an “elongated cluster of single-sex flowers bearing scaly bracts and usually lacking petals.”ref Male catkins appear on oak, chestnut, alder, birch, hazel, poplar, aspen, hornbeam, walnut and willow – a good overview of these is on the UK Tree Guide website and some examples are shown below. There are female catkins as well, but these don’t contain pollen, which is one way to tell them apart.
Some trees have flowers that look more like what we learn about at school, so-called ‘perfect’ flowers, with both male and female parts. Examples of trees with this type of flower structure include maples, hawthorn, lime, horse chestnut and many fruit trees including apple, cherry, pear and plum (see below).
Gymnosperms do not have flowers at all, instead they have male and female cones (also called strobili). 98% of gymnosperms use the wind for pollination, with the male pollen cones releasing their pollen into the wind to find its way to females.ref Because this is a bit of a hit and miss approach, gymnosperms produce a *lot* of pollen – one study estimated Juniper pollen production to be up to 532 billion pollen grains per tree!ref These vast quantities of pollen create pollen clouds, which you can see in a video on youtube here.
Another nifty thing is that female conifer cones produce a ‘pollination drop’ – this is a drop of liquid which sits on the surface of the cone to catch wind-borne pollen. As soon as pollination takes place, the drop is quickly retracted back into the cone.ref And you’ll notice that often male pollen cones are positioned at the top of a tree and in open positions along the stem (ie. not covered by leaves), to give their pollen the best chance of going far and wide.ref
Male cones on gymnosperms are smaller and less conspicuous than females, but, I think they’re still beautiful and worthy of our appreciation. Here are some examples of gymnosperm male cones:
Conifer species which have leaflets have a sort of tree-within-a-tree approach – such as the cypress below. – it has browny-pink male pollen cones on the ends of the shoots towards the back of the leaflet, and green developing female cones on the ends of the shoots at the tip of the leaflet.
There are more great images including scanning electron microscope images of gymnosperm pollen in this research paper.
But what if the trees in your bonsai collection don’t have any flowers or cones at all? Unfortunately this is a sign that they haven’t yet reached their reproductive phase. It’s hypothesised that a plant’s transition to its reproductive phase happens after a certain number of cell divisions have taken place.ref If you keep pruning the new growth off, your tree may never reach a reproductive phase, since it may never achieve the number of cell divisions required. The only way around this is to let your tree grow, use a mature tree to begin with, or use grafted material which comes from mature trees.
In the meantime, make sure to take a good look at your trees and the trees around you, and appreciate the underrated male tree flowers when they make an appearance.
Below is a shopping list if you’re wanting to create a bonsai pinetum. You may want to explore alternative species to include, if so the Gymnosperm database is a fantastic resource.
ARAUCARIACEAE
Monkey puzzle tree (Araucaria araucana) and/or Wollemi pine (Wollemia nobilis and/or Kauri (Agathis australis)
CUPRESSACEAE
Basic (5 genera): Hinoki cypress (Chamaecyparis obtusa), Dawn redwood (Metasequoia glyptostroboides), Japanese cedar/sugi (Cryptomeria japonica), Sabina Juniper (Juniperus sabina), and Thuja (eg. Thuja occidentalis)
Intermediate (10 genera): as above plus Giant redwood (Sequoiadendrongiganteum), Coast redwood ( Sequoia sempervirens), Swamp cypress (Taxodium distichum), Italian/Mediterranean cypress (Cupressus sempervirens) and an Oriental arborvitae (Platycladus orientalis).
Extensive (17 genera): as above plus Chinese Coffin Tree (Taiwaniacryptomerioides), Tasmanian Cedar/Pencil Pine or King Billy Pine (Athrotaxis cuppresoides/Athrotaxis selaginoides), Rottnest Island Pine or Oyster Bay Pine (Callitris preisii, Callitris rhomboidea), Chinese Fir (Cunninghamnia lanceolata) , Chilean Cedar(Austrocedrus chilensis), Incense Cedar (Calocedrus decurrens) and Chinese Swamp Cypress (Glyptostrobus pensilis)
Complete (25 genera): as above plus Diselma, Fitzroya, Libocedrus, Microbiota, Papuacedrus, Tetraclinis, Thujopsis and Widdringtonia.
PINACEAE
Basic (5 genera): Scot’s or Japanese Black or Japanese White Pine (Pinussylvestris/thunbergii/parviflora), Abies koreana (Korean fir), Cedrus atlantica (Atlantic cedar), Engelmann spruce (Picea engelmannii), European Larch (Larixdecidua)
Extensive (9 genera plus extras): as above plus Eastern white pine (Pinus strobus), Mountain Hemlock (Tsugamertensiana), Golden larch (Pseudolarix amabilis), Douglas fir (Pseudotsuga menziesii) and Yunnan youshan (Keteleeria evelyniana). Consider also Pinyon pine (Pinus monophylla)
Complete: all 11 genera: as above plus Cathay silver fir (Cathaya argyrophylla) and Bristlecone hemlock (Nothotsuga longibracteata)
PODOCARPACEAE
Basic: Buddhist Pine (Podocarpus macrophylla)
Extended: as above plus Celery Top Pine (Phyllocladus alpinus or Phyllocladus asplectiifolius), Chilean plum yew (Prumnopitys andina),
+ if you are a true collector and willing to track down seeds or specimens in collections near you, try for a Kahikatea (Dacrycarpus dacrydioides), Creeping Strawberry Pine (Microcachrys tetragona) and Rimu (Dacrydiumcupressinum)
SCIADOPITYACEAE
Japanese Umbrella Pine (Sciadopitysverticillata)
TAXACEAE
Basic: Common Yew (Taxus baccata) or Japanese Yew (Taxus cuspidata),
Intermediate: as above plus Japanese Plum Yew (Cephalotaxus harringtonii)
Extended: as above plus White Berry Yew (Pseudotaxus chienii) and Japanese Nutmeg Yew (Torreya nucifera)
+ for collectors Stinking Cedar (Torreya taxifolia)
A pinetum is an arboretum, or collection of trees, dedicated to conifers. There is a fabulous pinetum at RHS Wisley in the UK (my ‘About Me’ pic was taken there), and the UK National Pinetum at Bedgebury Forest has a collection of 12,000 specimen trees.
Whilst very few of us have the space to create a full-sized pinetum, the wonderful thing about bonsai is that you can create your own miniature version. There are only six conifer families, and within those, 68 genera, some of which would be impossible or at least extremely difficult to procure. So you could have a very respectable and representative bonsai pinetum with around 50 trees. A mame-sized bonsai pinetum might even fit on a single table!
Only a small number of conifer species are common bonsai subjects, so embarking on this project would require some creativity – there won’t be online tutorials or examples for many of these species. Some may be completely hopeless for bonsai (most of the Araucariaceae family for example), others may require conditions that you just can’t provide, but along the way I’m sure you would find a few that make excellent bonsai and give you something unique and different for your collection.
If you want to jump straight to the shopping list here it is, otherwise read on to find out about the trees in the list and where they fit in the different conifer families. For beginners to taxonomy, you start with a family, then a genus (or genera if there is more than one genus), then a species. So for example for Scot’s Pine Pinus sylvestris, Pinaceae is the family, Pinus is the genus, Sylvestris is the species.
You have a few different options for your representative tree/s from Araucariaceae as it has three genera (agathia, araucaria & wollemia) which cover a range of different forms. The most well-known in Europe would be the monkey puzzle tree (Araucaria araucana) but you could also include a Wollemi pine (Wollemia nobilis) which are available to buy, albeit at a cost. Neither of these are the easiest of bonsai subjects as they have a very regimented architecture with whorled branches, however they do have the advantage of being frost hardy. Wollemia nobilis also does backbud, and grows as a multi-stem.
An alternative could be an Agathis, also known as a Kauri tree. The New Zealand Agathis australis is the third largest known conifer after the giant and coast redwoodsref, depending on where it has been grown it may or may not be frost hardy. The New Zealand Bonsai Association has a Kauri forest on their native species web page.
If you live in the southern hemisphere, many of these options will be easier to find and will be happy outside.
The Cupressaceae conifer family is a lot easier to cover in your pinetum as it contains 25 genera (listed below) and 152 species.ref You can read about the leaves of many Cupressaceae species in my post on conifer scale leaves. It would be appropriate to include several members of this large family, as many are known as bonsai subjects anyway. An easy selection of five from different genera could include a Hinoki cypress (Chamaecyparis obtusa), a Dawn Redwood (Metasequoia glyptostroboides), a Japanese Cedar/Sugi (Cryptomeria japonica), a Sabina Juniper (Juniperus sabina), and a Thuja (also called arborvitae or cedars, although they are not true cedars).
Hinoki Cypress at RHS WisleyDawn Redwood ForestSugi/Cryptomeria Japonica at RHS WisleyTerry’s Sabina JuniperThuja occidentalis
Expanding to ten specimens across ten genera would allow the addition of five other reasonably easy to procure and grow species: Giant Redwood (Sequoiadendrongiganteum), Coast Redwood ( Sequoia sempervirens), Swamp Cypress (Taxodium distichum), Italian/Mediterranean Cypress (Cupressus sempervirens) and an Oriental Arborvitae (Platycladus orientalis).
If you’re a purist and want to include more genera from Cupressaceae,some excellent options would be the Chinese Coffin Tree (Taiwaniacryptomerioides), Tasmanian Cedar/Pencil Pine or King Billy Pine (Athrotaxis cuppresoides/Athrotaxis selaginoides), one of the Australian Callitris species such as Rottnest Island Pine or Oyster Bay Pine (Callitris preisii, Callitris rhomboidea), Chinese Fir (Cunninghamnia lanceolata) , Chilean Cedar (Austrocedrus chilensis), Incense Cedar (Calocedrus decurrens) and the Chinese Swamp Cypress (Glyptostrobus pensilis). You may need to grow these from seed, depending on where you live.
To fully represent Cupressaceae you’d also need to add the remaining eight genera, Diselma, Fitzroya, Libocedrus, Microbiota, Papuacedrus, Tetraclinis, Thujopsis and Widdringtonia. Many of these are specific to small or remote locations and/or endangered, but you may come across them while travelling, or while visiting full-sized pinetums or botanic gardens.
Family 3: Pinaceae
This family is a stalwart of the bonsai hobby, containing a massive (for conifers) 11 genera and 232 different species, including the eponymous pines, which account for more than half of these. You can read about the leaves of most Pinaceae in my post on conifer needle leaves. For your first pinetum Pinaceae, consider the UK native Scot’s Pine (Pinus sylvestris), Japanese black pine (Pinus thunbergii) or Japanese white pine (Pinus parviflora) – and perhaps a Pinus strobus which is a separate subgenus within Pinus.
For something unique, you could also include the only single-needled pine, Single-leaf piñon or Pinus monophylla. It’s actually a pine which has fused needles (five of them) which appear as one, and this results in its needles being very fat.
A top 5 selection from Pinaceae would also include species from the fir (Abies), true cedar (Cedrus), spruce (Picea) and larch (Larix) genera, all of which have species which are relatively easy to source and grow, at least in Europe (and most are extremely frost hardy). Let’s make it top 6 to include another well-known genus, the hemlock (Tsuga).
The 7th and 8th Pinaceae specimens could be a beautiful Golden larch (Pseudolarix amabilis the only species in its genus), and the classic Pacific north-west representative the Douglas fir (Pseudotsuga menziesii). The Douglas fir is not actually a fir, it’s a ‘false hemlock’, one of seven species of Pseudotsuga.
Most of these trees are well-known, can be procured either as plants or seeds, and could be relatively easily added to your collection. But the Pinaceae family also includes some extremely rare genera. Cathaya has only one species, Cathaya argyrophylla, or Cathay silver fir, which has a similar history to the famous Dawn redwood – thought to be a fossil but then ‘discovered’ in a small living stand in China in 1946.refCathaya was also discovered in China in 1938, but the discovery was not recognised as a new species until the 1950s.ref It is endangered with less than 1000 mature individuals in its native habitat.ref
Bristlecone hemlock or Nothotsuga is also a genus with only one species (Nothotsuga longibracteata) which comes from China, where it is near-threatened and “populations are highly fragmented, with some consisting of just a few scattered individuals”.ref Unfortunately Cathaya and Nothotsuga are probably out of reach for your pinetum unless you have access to seeds via a botanic garden, or live in China.
The 11th and final genus in Pinaceae is also relatively unknown outside of its native region of China, Taiwan & Vietnam – Keteleeria or ‘Yunnan youshan’ has three species, of which Keteleeriaevelyniana can be found as seed. So with Keteleeria evelyniana you can still have an unusual Pinaceae in your collection without having to raid your nearest full-sized pinetum.
Podocarpaceae is another large family within the conifers, with 172 species across 20 genera – so it is larger than Cupressaceae in terms of size but less well known in Europe and North America.ref This may be because podocarps are mainly found in tropical and subtropical mountain habitatsref, which has resulted in their leaves being quite different to other conifers (read more in my post on conifer flat leaves). It also results in trees from Podocarpaceae being a little harder to obtain in Europe.
There are three main groups within Podocarpaceae which could be represented in your pinetum – these are known as the ‘prumnopityoid clade’, the ‘dacrydioid clade’ and the ‘podocarpoid clade’.
The first group includes Phyllocladus – the so-called celery pines which come from Australasia. A somewhat hardy species includes Phyllocladus alpinus which I note can be purchased in the UK from Bluebell Nurseries, and seeds for other species such as Phyllocladus aspleniifolius are also available online. Depending on where you live these will need protection from hard frosts. Phylloclade species don’t have true leaves, aside from very small, almost invisible ones when they are seedlings. Instead they have ‘phylloclades’ which are photosynthetic flattened stemsref. So, of course you must have one of these interesting plants in your collection!
Also in the first group is the Prumnopitys genus from Polynesia and South America. Prumnopitys andina is known as the Chilean plum yew, and was the International Dendrology Society’s Tree of the Year in 2017. They produced a comprehensive report about this tree which you can read online, and which lists locations where they have been planted outside of Chile. It appears to be seasonally available in a small number of plant nurseries in the UK. I would have included a photo but no decent creative commons images were available, so I’ll add one when I get the chance to find a Chilean plum yew for myself.
The second group (the dacrydioid clade) includes the Dacrycarpus genus, of which Dacrycarpus dacrydioides is a species known and loved in New Zealand as ‘Kahikatea’ from the Maori. It’s the tallest tree in New Zealand, and apparently has a hardy form which can be bought from this provider in the UK. Below is one as a bonsai from the New Zealand bonsai association:
Also in this image is another member of the dacrydioid clade – which happens to be (according to Farjon) the smallest known conifer in the world. Microcachrys tetragona, or the Strawberry Pine, is from Tasmania. Bonsai enthusiast Diana Jones explains in the Newsletter of the Australian Plants as Bonsai Study Group: “Growing on the top of Mt. Wellington is a large tree, about 10m in diameter, called Microcachrys tetragona or creeping strawberry pine. Few people notice it because it is only about 5cm high, being negatively geotropic. This causes a few problems when making it into an attractive bonsai, because in a pot, it just flops.” Nevertheless I think her specimen has a certain charm, and she is definitely one tree ahead of me in the bonsai pinetum stakes.
And one can’t leave this group without mentioning Dacrydiumcupressinum, also from New Zealand and called variously red pine, red spruce or ‘Rimu’. My favourite podcaster has covered this species on his blog In Defense of Plants where he says the fleshy cones of this tree are an essential part of the diet of the endangered kākāpō bird. Availability of the ‘fruit’ (not really since it’s a conifer – it’s a female cone) triggers breeding for the kākāpō so it’s seen as critical to their survival.ref The tree itself is a wonderful tree with long dangly stems so I’d love to have one even without a kākāpō.
But if you cannot find Kahikatea, Tasmanian creeping strawberry pine or a kākāpō-infested Rimu, the third group in Podocarpaceae (the Podocarpoid clade) is going to be much easier to represent in your pinetum, because it contains the oft-seen bonsai species the Buddhist pine, or Podocarpus macrophyllus (also called Kusamaki in Japanese). If you are going to have at least one specimen Podocarp, this is likely to be the easiest one to obtain. I have a couple in my London garden (in the ground) but after the last brutal winter I think they would prefer to be indoors, and they are sold as indoor bonsai in the UK.
There are various other Podocarpus species available at nurseries such as Podocarpus salignus or willow-leaved podocarp – it’s probably a good idea to do a local search to see which species are available in your area.
Sciadopityaceae has only one genus (Sciadopitys) and within that only one species, the Japanese umbrella pine or Sciadopitysverticillata. This tree is an endemic Japanese evergreen conifer, with relatively slow growth rates, used in gardens and construction in Japan.ref1,ref2
Unfortunately since this is the only representative of one of the six conifer families, you really do need one in your collection if you are to truly represent all the conifers. This tree isn’t very easy to propagate, and is also quite expensive to buy (at least in the UK), not only that, it doesn’t appear very commonly as a bonsai. I haven’t had any luck growing it from seed (and they were expensive) so I think your best best is keeping an eye on nurseries and waiting until you see one at a reasonable price.
Finally we come to the yews, the Taxaceae family which has six genera and 28 species. Many will be familiar with its most prominent genus, Taxus, containing the Common yew Taxus baccata. According to the gymnosperm database, most members of Taxus look pretty much the same, so to save yourself money and time, I’d suggest simply finding the yew that is easily available in your area. For me Taxus baccata are a dime a dozen, you also see Japanese Yew Taxus cuspidata used in bonsai.
I believe you should also have a Japanese plum yew in your collection. Cephalotaxus is a genus with 11 species, which used to be considered its own family, but DNA testing revealed it really belonged in Taxaceae.ref This tree has yew-like leaves and small plum-like fleshy cones which start green and then move through red and dark purple colourationref. It’s not commonly seen as a bonsai but it’s not difficult to propagate and I have seen them available at plant nurseries in the UK. One would assume adopting a similar styling approach to yew would work with this tree.
The remaining genera in Taxaceae are a lot rarer and more difficult to include in any non-tropical pinetum. The New Caledonia Yew Austrataxus spicata is the only southern hemisphere Taxaceae, and thrives in the very unusual habitat of the island, based on ultramafic rock containing chrome and nickel and not much else in the way of nutrients.ref
The Catkin Yew Amentotaxus is a threatened genus with six species found in China, India, Laos, Vietnam and Taiwan.ref One nursery in the UK sells Amentotaxus argotaenia var. argotaenia as a pot-grown specimen to be taken indoors during winter (but when I checked they were out of stock).
The White Berry Yew, Pseudotaxus chienii, is the only species in the Pseudotaxus genus, and is also native to China.ref It has white arils instead of the red arils of Taxus baccata. Cited as rare, it is nevertheless available from some suppliers.
The final genus of Taxaceae and of this pinetum article, is a really interesting one called Torreya which I think deserves a place representing this family alongside the more familiar trees mentioned above.
Torreya nucifera was the International Dendrology Society’s Tree of the Year (2019) and has a full report write-up available online here. This tree is known as the Nutmeg Yew, or in Japanese ‘kaja’ and ‘kaya’ and oil from its seeds (not nuts!) have been used for tempura cooking oil. As a widely cultivated tree, there is availability of Torreya from plant nurseries (eg. here) although the article I linked to says they do not like cool summers so a protected position or a pot may be needed.
Another option from Torreya if you can find one, and have the budget, is the alarmingly named Stinking Cedar or Torreya taxifolia. This is a rare species native to Florida USA, now protected in the Torreya State Park in Florida. One can be acquired for £50 per 2L pot in the UK here.
The end (of this post, but the start of your conifer collection?)
So there you have it, a set of suggestions for creating your own mini-pinetum using bonsai trees across the six conifer families. If you decide to take on the challenge, I’d love to see your efforts – tag me on Facebook (Bonsai-Science) or twitter (@BonsaiScience). Here’s another link to the shopping list, to get you started.